Водоросли — общая характеристика. Классификация, питание, места обитания. Особенности строения, процессов жизнедеятельности и распространение водорослей Строение и значение водорослей
Понятие «водоросли» в научном отношении неопределенно. Слово «водоросли» буквально означает лишь то, что это растения, живущие в воде, однако не все растения в водоемах можно с научной точки зрения назвать водорослями, такие растения, как тростник, камыш, рогоз, кувшинки, кубышки, мелкие зеленые пластинки ряски и др., являются семенными (или цветковыми) растениями. К этим растениям научный термин «водоросли» неприменим, их называют водяными растениями
Понятие «водоросли» – не систематическое, а биологическое. Водоросли (Algae ) – это сборная группа организмов, основная часть которых, согласно современным представлениям, входит в царство Растений (Plantae ), в котором она составляет два подцарства: багрянки, или красные водоросли – Rhodobionta и настоящие водоросли – Phycobionta (в третье подцарство царства Растений входят высшие (зародышевые или листостебельные) растения – Embryobionta ). Остальные организмы, относимые к водорослям, сейчас уже не считаются растениями: синезеленые и прохлорофитовые водоросли часто считают самостоятельной группой или относят к бактериям, а эвгленовые водоросли иногда относят к подцарству животных – простейшим. Различные группы водорослей возникли в разное время и, по-видимому, от разных предков, но в результате эволюции в сходных условиях обитания приобрели много сходных черт.
У организмов, объединяемых в группу водорослей, есть ряд общих признаков. В морфологическом отношении для водорослей наиболее существенным признаком является отсутствие многоклеточных органов – корня, листьев, стебля, типичных для высших растений. Такое нерасчлененное на органы тело водорослей носит название слоевище, или таллом.
У водорослей более простое (по сравнению с высшими растениями) анатомическое строение – нет проводящей (сосудистой) системы, поэтому водоросли, относимые к растениям, являются бессосудистыми растениями. Водоросли никогда не образуют цветков и семян, а размножаются вегетативно или спорами.
В клетках водорослей содержится хлорофилл, благодаря которому они способны ассимилировать на свету углекислый газ (т.е. питаться при помощи фотосинтеза), это преимущественно обитатели водной среды, но многие приспособились к жизни в почве и на ее поверхности, на скалах, на стволах деревьев и в других биотопах.
Организмы, относимые к водорослям чрезвычайно разнородны. Водоросли принадлежат как к прокариотам (доядерным организмам), так и к эукариотам (истинно ядерным организмам). Тело водорослей может быть всех четырех степеней сложности, вообще известных для организмов: одноклеточным, колониальным, многоклеточным и неклеточным, размеры их колеблются в очень широких пределах: мельчайшие соизмеримы с бактериальными клетками (не превышают 1 мкм в диаметре), а наиболее крупные морские бурые водоросли достигают 30–45 м в длину.
Водоросли разделяются на большое количество отделов и классов и разделение их на систематические группы (таксоны) производится по биохимическим особенностям (набор пигментов, состав клеточной оболочки, тип запасных веществ), а также по субмикроскопическому строению. Однако для современной систематики водорослей характерно множество разнообразных систем. Даже на самых высоких таксономических уровнях (надцарства, подцарства, отделы и классы) систематики не могут прийти к единому мнению.
По одной из современных систем, водоросли делятся на 12 отделов: синезеленые, прохлорофитовые, красные, золотистые, диатомовые, криптофитовые, динофитовые, бурые, желтозеленые, эвгленовые, зеленые, харовые. Всего известно около 30 тысяч видов водорослей.
Наука о водорослях называется альгология или фикология, ее рассматривают как самостоятельный раздел ботаники . Водоросли являются объектами для решения вопросов, относящихся к другим наукам (биохимии , биофизики , генетики и др.) Данные альгологии учитывают при разработке общебиологических проблем и хозяйственных задач. Развитие прикладной альгологии идет в трех основных направлениях: 1) использование водорослей в медицине и в различных областях хозяйства; 2) для решения природоохранных вопросов; 3) накопление данных о водорослях для решения задач других отраслей.
Строение водорослей.
Основной структурной единицей тела водорослей, представленных одноклеточными и многоклеточными формами, является клетка. Существуют различные типы клеток водорослей, их разделяют по форме (шаровидные, цилиндрические и т. д.), функциям (половые, вегетативные, способные и не способные к фотосинтезу и др.), расположению и пр. Но наиболее принципиальной в наши дни считается классификация клеток по особенностям их тонкого строения, обнаруживаемого с помощью электронного микроскопа. С этой точки зрения различают клетки, содержащие типичные ядра, (т.е. ядра, окруженные ядерными оболочками, мембранами), и клетки, не имеющие типичных ядер. Первый случай – эукариотическое строение клетки, втором – о прокариотическое. Прокариотическое строение клетки имеют синезеленые и прохлорофитовые водоросли, эукариотическое – представители всех других отделов водорослей.
Вегетативное тело водорослей (слоевище) отличается морфологическим разнообразием, водоросли могут быть одноклеточными, колониальными, многоклеточными и неклеточными. Размеры их в пределах каждых из этих форм колеблются в широких пределах – от микроскопических до очень крупных.
Особенность одноклеточных форм водорослей определяется тем, что их организм состоит из одной клетки, поэтому в ее строении и физиологии сочетаются клеточные и организменные черты. Это автономная система, способная расти и самовоспроизводиться, мелкая, не видимая простым глазом одноклеточная водоросль является своеобразной фабрикой, которая добывает сырье (поглощая из окружающей среды растворы минеральных солей и углекислоты), перерабатывает и производит такие ценные соединения, как белки, углеводы и жиры. Кроме того, важными продуктами ее жизнедеятельности является кислород и углекислый газ и, таким образом, она активно участвует в круговороте веществ в природе. Одноклеточные водоросли иногда образуют временные или постоянные скопления (колонии).
Многоклеточные формы возникли после того, как клетка проделала длительный и сложный путь развития как самостоятельный организм. Переход от одноклеточного к многоклеточному состоянию сопровождался потерей индивидуальности и связанными с этим изменениями в структуре и функциях клетки. Внутри талломов многоклеточных водорослей складываются качественно иные отношения, чем между клетками одноклеточных водорослей. С возникновением многоклеточности появилась дифференциация и специализация клеток в талломе. С эволюционной позиции это следует рассматривать как первый шаг на пути становления тканей и органов.
Уникальную группу составляют сифоновые водоросли: у них талломы не поделены на клетки, однако и у них в цикле развития есть одноклеточные стадии.
Окраска водорослей многообразна (зеленая, розовая, красная, оранжевая, почти черная, фиолетовая, голубая и др), обусловлено это тем, что одни водоросли содержат только хлорофилл, а другие – еще ряд пигментов, окрашивающих их в различные цвета.
Водоросли (а точнее, синезеленые водоросли, или цианобактерии) были первыми организмами на Земле, у которых в процессе эволюции появилась способность к фотосинтезу, процессу образования органических веществ под воздействием света. Как источник углерода при фотосинтезе используется углекислый газ (CO 2), в качестве источника водорода – вода (H 2 O), а в результате выделяется свободный кислород.
Тип питания с помощью фотосинтеза, при котором организм, используя энергию фотосинтеза, синтезирует все необходимые органические вещества из неорганических, стал одним из основных способов питания водорослей и других зеленых растений. Однако многие водоросли могут при определенных условиях достаточно легко переключаться с фотосинтезного способа питания на ассимиляцию различных органических соединений, при этом организм использует для питания готовые органические вещества, или сочетает этот способ питания с фотосинтезом.
Помимо использования органических соединений как источника углерода, водоросли могут переключаться с ассимиляции неорганического нитратного азота на усвоение азота из органических соединений, некоторые синезеленые водоросли могут вообще обходиться без связанных форм азота и фиксировать свободный азот из атмосферы как азотофиксирующие организмы.
Многообразие способов питания водорослей позволяют им иметь широкие ареалы и занимать разнообразные экологические ниши.
Воспроизведение себе подобных у водорослей совершается посредством вегетативного, бесполого и полового размножения .
Происхождение водорослей.
Вопрос о происхождении и эволюции водорослей очень сложен из-за разнообразия этих растений, особенно их субмикроскопического строения и биохимических признаков, кроме того, большинство водорослей в ископаемом состоянии не сохранилось и нет связующих звеньев между современными отделами растений в виде организмов промежуточного строения.
Проще всего решается вопрос о происхождении прокариотических (доядерных) водорослей – синезеленых, имеющих много общих признаков с фотосинтезирующими бактериями. Вероятнее всего, синезеленые водоросли произошли от организмов, близких к пурпурным бактериям и содержащих хлорофилл ().
О происхождении эукариотических (ядерных) водорослей сейчас нет единой точки зрения. Есть две группы теорий, исходящих или из симбиотического, или из несимбиотического их происхождения, однако против каждой из этих теорий есть свои возражения.
По теории симбиогенеза, хлоропласты и митохондрии клеток эукариотических организмов некогда были самостоятельными организмами: хлоропласты – прокариотическими водорослями, митохондрии – аэробными бактериями (). В результате захвата амебоидными эукариотическими организмами аэробных бактерий и прокариотических водорослей возникли предки современных групп эукариотических водорослей. Некоторые исследователи приписывают симбиотическое происхождение также хромосомам и жгутикам.
По теории несимбиотического происхождения эукариотические водоросли возникли от предка, общего с синезелеными водорослями, имеющего хлорофилл и фотосинтез с выделением кислорода, в этом случае современные фотосинтезирующие прокариоты (синезеленые водоросли) – это боковая, тупиковая ветвь растительной эволюции.
Основные факторы, влияющие на развитие водорослей.
Основными факторами, влияющими на развитие водорослей, являются свет, температура, наличие воды, источников углерода, минеральных и органических веществ. Водоросли широко распространены по всему земному шару, их можно встретить в воде, в почве и на ее поверхности, на коре деревьев, стенах деревянных и каменных построек и даже в таких негостеприимных местах, как пустыни и ледники.
Факторы, влияющие на развитие водорослей, подразделяют на абиотические, не связанные с деятельностью живых организмов, и биотические, обусловленные этой деятельностью. Многие факторы, особенно абиотические, являются лимитирующими, т.е. они способны ограничивать развитие водорослей. Жизнь всех организмов, в том числе и водорослей, зависит от содержания в среде обитания необходимых веществ, значения физических факторов, а также диапазона устойчивости самих организмов относительно изменений условий среды. Уровень, при котором конкретный фактор может выступать как лимитирующий, для разных видов водорослей различен. В водных экосистемах к лимитирующим факторам относятся температура, прозрачность, наличие течения, концентрация кислорода, углекислого газа, солей и биогенных веществ. В наземных местообитаниях основными лимитирующими факторами являются климатические: температура, влажность, свет и т.д., а также состав и строение субстрата. Эти две группы факторов вместе с популяционными взаимодействиями определяют характер наземных сообществ и экосистем.
Для большинства водорослей вода – постоянная среда обитания, однако многие их виды могут жить и вне воды. Среди растений, обитающих на суше, по устойчивости к высыханию выделяют пойкилогидрические, не способные поддерживать постоянное содержание воды в тканях, и гомойгидрические, способные поддерживать постоянную гидратацию тканей. У пойкилогидрических водорослей (синезеленые и некоторые зеленые водоросли) клетки при высыхании сжимаются без необратимого изменения ультраструктуры и, следовательно, не теряют жизнеспособности, при увлажнении у них восстанавливается нормальный обмен веществ. Минимальная влажность, при которой возможна нормальная деятельность таких растений, различна. Клетки гомойгидрических водорослей при высыхании погибают, поэтому такие растения, как правило, обитают при постоянно избыточной увлажненности. К гомойгидрическим водорослям относятся, например, некоторые виды зеленых и желтозеленых водорослей.
Соленость и минеральный состав воды – это важнейшие лимитирующие факторы, влияющие на распределение водорослей.
Водоросли обитают в водоемах самой различной солености: от пресных водоемов, минерализация которых не превышает обычно 0,5 г/л, до крайне засоленных (гипергалинных) водоемов, концентрация солей которых находится в пределах от 40 до 347 г/л. Несмотря на то, что в целом водорослям свойственна такая широкая амплитуда солеустойчивости, конкретные виды в большинстве своем стеногалинные , т.е. способны обитать лишь при определенном значении солености. Эвригалинных видов водорослей, способных существовать при разной солености, сравнительно немного.
Кислотность воды также является лимитирующим фактором. Устойчивость разных таксонов водорослей к изменениям кислотности (рН) столь же различна, как и к изменениям солености. Некоторые виды водорослей живут только в щелочных водах, при высоком значении рН, другие обитают в кислых водах, при низком рН.
Наличие в среде макро- и микроэлементов, являющихся необходимыми компонентами тела водорослей, имеет решающее значение для интенсивности их развития.
Элементы и их соединения, относящиеся к макроэлементам, требуются организмам в сравнительно больших количествах. Наиболее важны азот и фосфор, почти столь же необходимы калий, кальций, сера и магний.
Микроэлементы необходимы растениям в крайне малых количествах, но они имеют огромное значение для их жизни, поскольку входят в состав многих жизненно-важных ферментов. Микроэлементы нередко выступают как лимитирующие факторы. К ним относятся 10 элементов: железо, марганец, цинк, медь, бор, кремний, молибден, хлор, ванадий и кобальт.
Водоросли разных отделов имеют неодинаковые потребности в макро- и микроэлементах. Например, для нормального развития диатомовых водорослей требуется довольно значительное количество кремния, который используется для постройки их панциря. При недостатке кремния панцири диатомовых истончаются.
Почти во всех пресноводных и морских экосистемах лимитирующим фактором является концентрация в воде нитратов и фосфатов. В пресных водоемах с низким содержанием карбонатов к лимитирующим факторам могут быть причислены концентрации солей кальция и некоторых других.
Свет необходим водорослям как источник энергии фотохимических реакций и как регулятор развития. Его избыток, также как и недостаток, может быть причиной серьезных нарушений развития водорослей. Поэтому свет тоже является лимитирующим фактором при слишком большой или слишком маленькой освещенности.
Распределение водорослей в толще воды в значительной степени определяется наличием света, необходимого для нормального фотосинтеза. Слой воды выше границы обитания фотоавтотрофных организмов называется эвфотической зоной . В море граница эвфотической зоны обычно находится на глубине 60 м, изредка опускаясь до глубины 120 м, а в прозрачных водах океана – приблизительно до 140 м. В озерных, значительно менее прозрачных водах, граница этой зоны обычно проходит на глубине 10–15 м, а в наиболее прозрачных гляциальных и карстовых озерах – на глубине 20–30 м.
Оптимальные значения освещенности для разных видов водорослей варьируются в широких пределах. По отношению к свету выделяют гелиофильные и гелиофобные водоросли. Гелиофильные (светолюбивые) водоросли для нормальной жизнедеятельности нуждаются в значительном количестве света. К ним относится большинство синезеленых и значительное количество зеленых водорослей, обильно развивающихся в летнее время в поверхностных слоях воды. Гелиофобные (избегающие яркого света) водоросли приспособлены к условиям низкой освещенности. Например, большинство диатомовых водорослей избегает ярко освещенного поверхностного слоя воды и в малопрозрачных водах озер интенсивно развивается на глубине 2–3 м, а в прозрачных водах морей – на глубине 10–15 м.
У водорослей разных отделов в зависимости от состава особых светочувствительных пигментов, максимальная активность фотосинтеза наблюдается при разной длине световых волн. В наземных условиях частотные характеристики света довольно постоянны, поэтому постоянна и интенсивность фотосинтеза. При прохождении через воду свет красной и синей области спектра поглощается, и на глубину проникает зеленоватый свет, слабо воспринимаемый хлорофиллом. Поэтому там выживают в основном красные и бурые водоросли, имеющие дополнительные фотосинтезирующие пигменты, способные использовать энергию зеленого света. Отсюда становится понятным огромное влияние света на вертикальное распределение водорослей в морях и океанах: в приповерхностных слоях, как правило, преобладают зеленые водоросли, глубже – бурые, и на наиболее глубоководных участках – красные. Однако подобная закономерность не является абсолютной. Многие водоросли способны существовать в условиях крайне низкой, не свойственной им освещенности, а иногда и в полной темноте. При этом у них могут происходить определенные изменения в пигментном составе или в способе питания. Так, представители многих отделов водорослей способны при отсутствии света и избытке органических веществ переходить к питанию органическими соединениями мертвых тел или экскрементов животных.
Для водорослей, обитающих в водных биотопах, движение воды играет огромную роль. Движение водных масс обеспечивает приток питательных веществ и удаление продуктов жизнедеятельности водорослей. В любых континентальных и морских водоемах существует относительное движение водных масс, поэтому почти все водоросли водоемов – обитатели текучих вод. Исключение составляют лишь водоросли, развивающиеся в особо крайних условиях (в пустотах скал, толще льда и пр.).
Водорослям свойственны очень широкие диапазоны температурной устойчивости. Некоторые их виды способны существовать как в горячих источниках, температура которых близка к температуре кипения воды, так и на поверхности льда и снега, где температуры колеблются около 0° С.
По отношению к температуре среди водорослей выделяют: эвритермные виды , существующие в широком температурном диапазоне (например, зеленые водоросли из порядка Oedogoniales, стерильные нити которых можно найти в мелких водоемах с ранней весны до поздней осени), и стенотермные , приспособленные к очень узким, иногда экстремальным температурным зонам. К стенотермным относятся, например, криофильные (холодолюбивые) водоросли, растущие только при температуре, близкой к 0° С и термофильные (теплолюбивые) водоросли, не способные существовать при температурах ниже 30° С.
Температура определяет географическое распространение водорослей, развивающихся в водной среде. В целом, за исключением широко распространенных эвритермных видов, в распространении водорослей наблюдается географическая зональность: конкретные таксоны морских планктонных и бентосных водорослей приурочены к определенным географическим поясам. Так, крупные бурые водоросли (Macrocystis) доминируют в северных морях. По мере продвижения к югу все более заметную роль начинают играть красные водоросли, а бурые отходят на второй план. В фитопланктоне тропических вод чрезвычайно богато представлены динофитовые и золотистые водоросли. В северных морях в фитопланктоне господствуют диатомовые водоросли. Температура оказывает влияние и на вертикальное распределение планктонных и бентосных водорослей. Здесь она воздействует в основном косвенным образом, ускоряя или замедляя темпы роста отдельных видов, что приводит к их вытеснению другими видами, растущими более интенсивно в данном температурном режиме.
Водоросли, входя в состав экосистем, связаны с остальными их компонентами множественными связями. Претерпеваемые водорослями прямые и косвенные воздействия, обусловленные жизнедеятельностью других организмов, относят к биотическим факторам.
В большинстве случаев в экосистеме водоросли выступают как продуценты органического вещества. Поэтому важнейшим фактором, ограничивающим развитие водорослей в конкретной экосистеме, является наличие животных, существующих за счет поедания водорослей.
Различные виды водорослей способны оказывать воздействие друг на друга, путем выделения химических веществ во внешнюю среду (такое взаимодействие растений называется аллелопатией ). Иногда это является препятствием для их совместного существования.
У некоторых видов водорослей могут складываться конкурентные отношения друг с другом за места обитания.
Человек оказывает значительное воздействие на природные экосистемы, что делает антропогенный фактор весьма существенным для развития водорослей. Прокладывая каналы и сооружая водохранилища, человек создает новые местообитания для водных организмов, нередко принципиально отличающиеся от водоемов данного региона по гидрологическому и тепловому режиму. Сбросы сточных вод нередко приводят к обеднению видового состава и гибели водорослей или к массовому развитию отдельных видов. Первое происходит при сбросе токсичных вод, второе – при обогащении водоема биогенными веществами (особенно соединениями азота и фосфора). Следствием неумеренного сброса биогенных веществ в водоем может быть его эвтрофикация, которая приводит к бурному развитию водорослей («цветение воды»), дефициту кислорода, замору рыб и других водных животных. На водоросли, особенно аэрофитные и почвенные, могут оказывать влияние и атмосферные выбросы токсичных промышленных отходов. Очень часто последствия вмешательства человека в жизнь экосистем имеют необратимый характер.
Экологические группы водорослей.
Водоросли распространены по всему земному шару и встречаются в различных водных, наземных и почвенных биотопах. Известны разнообразные экологические группы этих организмов: 1) планктонные водоросли; 2) нейстонные водоросли; 3) бентосные водоросли; 4) наземные водоросли; 5) почвенные водоросли; 6) водоросли горячих источников; 7) водоросли снега и льда; 8) водоросли соленых водоемов; 9) водоросли, существующие в известковом субстрате.
Водоросли водных местообитаний.
Планктонные водоросли.
Планктон – это совокупность организмов, населяющих толщу воды континентальных и морских водоемов и не способных противостоять переносу течениями (т.е. как бы парящих в воде). В состав планктона входят фито-, бактерио- и зоопланктон.
Фитопланктоном называют совокупность свободноплавающих в толще воды мелких, преимущественно микроскопических растений, основную массу которых составляют водоросли. Фитопланктон населяет только эвфотическую зону водоемов (поверхностный слой воды с достаточной для фотосинтеза освещенностью).
Планктонные водоросли обитают в самых разнообразных водоемах – от маленькой лужи до океана. Их нет лишь в водоемах с резко аномальным режимом, в том числе в термальных (при температуре воды выше +80° С и заморных (зараженных сероводородом) водоемах, в чистых приледниковых водах, не содержащих минеральных питательных веществ, а также в пещерных озерах. Суммарная биомасса фитопланктона невелика по сравнению с биомассой зоопланктона (соответственно 1,5 и более 20 млрд. т), но из-за быстрого размножения его продукция в Мировом Океане составляет около 550 млрд. тонн в год, что почти в 10 раз больше суммарной продукции всего животного населения океана.
Фитопланктон – основной продуцент органического вещества в водоемах, за счет которого существуют водные гетеротрофные животные и некоторые бактерии. Фитопланктон является начальным звеном большинства пищевых цепей в водоеме: им питаются мелкие планктонные животные, которыми питаются более крупные. Поэтому в районах наибольшего развития фитопланктона обильны зоопланктон и нектон.
Состав и экология отдельных представителей водорослевого фитопланктона в разных водоемах чрезвычайно разнообразны. Общее число видов фитопланктона во всех морских и внутренних водоемах достигает 3000.
Обилие и видовой состав фитопланктона зависит от комплекса рассмотренных выше факторов. В связи с этим видовой состав планктонных водорослей в разных водоемах (и даже в одном и том же водоеме, но в разное время года) не одинаков. Он зависит от физического и химического режима в водоеме. В каждый сезон года преобладающее развитие получает одна из групп водорослей (диатомовые, синезеленые, золотистые, эвгленовые, зеленые и некоторые другие), причем нередко господствует всего один вид той или иной группы. Особенно это выражено в пресноводных водоемах.
Во внутренних водоемах существует гораздо большее разнообразие экологических условий по сравнению с морскими водоемами, что определяет и значительно большее разнообразие видового состава и экологических комплексов пресноводного фитопланктона по сравнению с морским. Одной из существенных особенностей пресноводного фитопланктона является обилие в нем временно планктонных водорослей. Ряд видов, которые принято считать типично планктонными, в прудах и озерах имеют донную или перифитонную (прикрепление к какому-либо предмету) фазу в своем развитии.
Морской фитопланктон состоит в основном из диатомовых и динофитовых водорослей. Хотя морская среда на значительных пространствах относительно однородна, в распределении морского фитопланктона однородности не наблюдается. Различия по видовому составу и численности нередко выражены даже на сравнительно небольших акваториях морских вод, но особенно четко они отражаются в крупномасштабной географической зональности распределения. Здесь проявляется действие основных факторов среды: солености воды, температуры, освещенности и содержания питательных веществ.
Планктонные водоросли обычно имеют специальные приспособления к обитанию в толщи воды во взвешенном состоянии. У одних видов это разного рода выросты и придатки тела – шипы, щетинки, роговые отростки, перепонки, парашюты; другие образуют полые или плоские колонии и обильно выделяют слизь; третьи накапливают в своем теле вещества, удельный вес которых меньше удельного веса воды (капли жира у диатомовых и некоторых зеленых водорослей, газовые вакуоли у синезеленых). Эти образования гораздо сильнее развиты у морских фитопланктеров, чем у пресноводных. Еще одним из таких приспособлений являются мелкие размеры тела планктонных водорослей.
Нейстонные водоросли.
Совокупность морских и пресноводных организмов, обитающих у поверхностной пленки воды, прикрепляющихся к ней или передвигающихся по ней называется нейстоном. Нейстонные организмы обитают как в мелких водоемах (прудах, заполненных водой ямах, небольших заливах озер), так и в крупных, в том числе в морях. В отдельных случаях они развиваются в таком количестве, что покрывают воду сплошной пленкой.
В состав нейстона входят одноклеточные водоросли, входящие в состав разных систематических групп (золотистые, эвгленовые, зеленые, отдельные виды желтозеленых и диатомовых). Некоторые нейстонные водоросли имеют характерные приспособления для существования у поверхности воды (например, слизистые или чешуйчатые парашюты, удерживающие их на поверхностной пленке).
Бентосные водоросли.
К числу бентосных (донных) водорослей относятся водоросли, приспособленные к существованию в прикрепленном или неприкрепленном состоянии на дне водоемов и на разнообразных предметах, живых и мертвых организмах, находящихся в воде.
Преобладающими бентосными водорослями континентальных водоемов являются диатомовые, зеленые, синезеленые и желтозеленые многоклеточные (нитчатые) водоросли, прикрепленные или неприкрепленные к субстрату.
Основные бентосные водоросли морей и океанов – бурые и красные, иногда зеленые макроскопические прикрепленные слоевищные формы. Все они могут обрастать мелкими диатомовыми, синезелеными и другими водорослями.
Иногда водоросли, растущие на предметах, введенных в воду человеком (суда, плоты, буи) относят к перифитону . Выделение этой группы обосновывается тем, что входящие в ее состав организмы (водоросли и животные) живут на предметах движущихся или обтекаемых водой. Кроме того, эти организмы удалены от дна и, следовательно, находятся в условиях иного светового и температурного режимов, а также в других условиях поступления биогенных веществ.
Возможность произрастания бентосных водорослей в конкретных местообитаниях определяется как абиотическими, так и биотическими факторами. Среди последних существенную роль играет конкуренция с другими водорослями и присутствие животных, питающихся водорослями (морских ежей , брюхоногих моллюсков , ракообразных , рыб). Воздействие биотических факторов приводит к тому, что отдельные виды водорослей растут далеко не на всякой глубине и не во всяких водоемах с подходящим световым и гидрохимическим режимом.
К абиотическим факторам относятся свет, температура, а также содержание в воде биогенных и биологически активных веществ, кислорода и неорганических источников углерода. Очень важна скорость поступления этих веществ в слоевище, что находится в зависимости от концентрации веществ и скорости движения воды.
Бентосные водоросли, растущие в условиях движения воды, получают преимущества по сравнению с водорослями, растущими в малоподвижных водах. Один и тот же уровень фотосинтеза может быть достигнут у них при меньшей освещенности, что способствует росту более крупных слоевищ; движение воды предотвращает оседание на скалы и камни илистых частиц, которые мешают закреплению зачатков водорослей, а также смывает с поверхности грунта питающихся водорослями животных. К тому же, несмотря на то, что при сильном течении или сильном прибое происходит повреждение слоевищ водорослей или отрыв их от грунта, движение воды все же не препятствует поселению микроскопических водорослей и микроскопических стадий крупных водорослей. Поэтому места с интенсивным движением воды (в морях это проливы с течениями, прибрежные участки прибоя, в реках – камни на перекатах) отличаются пышным развитием бентосных водорослей.
Влияние движения воды на развитие бентосных водорослей особо ощутимо в реках, ручьях, горных потоках. В этих водоемах выделяется группа бентосных организмов, предпочитающих места с постоянным течением. В озерах, где не бывает сильных течений, основное значение приобретает волновое движение. В морях волны также оказывают значительное влияние на жизнь бентосных водорослей, в частности на их вертикальное распределение.
В северных морях на распространение и численность бентосных водорослей оказывает влияние лед. Заросли водорослей могут быть уничтожены (стерты) движением ледников. Поэтому, например, в Арктике многолетние водоросли легче всего найти у берега среди валунов и выступов скал, препятствующих движению льда.
Интенсивному развитию бентосных водорослей способствует также умеренное содержание в воде биогенных веществ. В пресных водах такие условия создаются в неглубоких прудах, в прибрежной зоне озер, в речных заводях, в морях – в мелких заливах. Если в таких местах существует достаточное освещение, твердые грунты и слабое движение воды, то создаются оптимальные условия для жизни фитобентоса. При отсутствии движения воды и ее недостаточном обогащении биогенными веществами, бентосные водоросли растут плохо.
Водоросли горячих источников.
Водоросли, выдерживающие высокие температуры, называются термофильными . В природе они поселяются в горячих источниках, гейзерах и вулканических озерах. Нередко они обитают в водах, которые кроме высокой температуры характеризуются повышенным содержанием солей или органических веществ (сильно загрязненные горячие сточные воды заводов, фабрик, электростанций или атомных станций).
Предельные температуры, при которых удавалось находить термофильные водоросли, судя по разным источникам, колеблются от 52 до 84° С. Всего обнаружено около 200 видов термофильных водорослей, однако видов, живущих только при высоких температурах, среди них сравнительно немного. Большинство из них способно выдерживать высокие температуры, но обильнее развиваются при обычных температурах. Типичными обитателями горячих вод являются синезеленые, в меньшей степени – диатомовые и некоторые зеленые водоросли.
Водоросли снега и льда.
Водоросли снега и льда составляют подавляющее большинство организмов, поселяющихся на замерзших субстратах (криобиотопах). Общее число видов водорослей, обнаруженных на криобиотопах, достигает 350, но истинных криофилов, способных вегетировать только при температурах, близких к 0° С, значительно меньше: немногим более 100 видов. Это микроскопические водоросли из которых подавляющее большинство относится к зеленым водорослям (около 100 видов); несколькими видами представлены синезеленые, желтозеленые, золотистые, пирофитовые и диатомовые водоросли. Все эти виды обитают в поверхностных слоях снега или льда. Их объединяет способность выдерживать замерзание без нарушения тонких клеточных структур и затем, при оттаивании, быстро возобновлять вегетацию, используя минимальное количество теплоты. Лишь немногие из них имеют стадии покоя, большинство лишены каких-либо специальных приспособлений для перенесения низких температур.
Развиваясь в массовом количестве, водоросли способны вызывать зеленое, желтое, голубое, красное, коричневое, бурое или черное «цветение» снега и льда.
Водоросли соленых водоемов.
Эти водоросли вегетируют при повышенной концентрации в воде солей, достигающей 285 г/л в озерах с преобладанием поваренной соли и 347 г/л в глауберовых (содовых) озерах. По мере увеличения солености количество видов водорослей уменьшается, очень высокую соленость переносят лишь немногие из них. В крайне засоленных (гипергалинных) водоемах преобладают одноклеточные подвижные зеленые водоросли. Нередко они вызывают красное или зеленое «цветение» соленых водоемов. Дно гипергалинных водоемов иногда сплошь покрыто синезелеными водорослями. они играют большую роль в жизни соленых водоемов. Сочетание органической массы, образуемой водорослями, и большого количества растворенных в воде солей обуславливает ряд своеобразных биохимических процессов, свойственных этим водоемам. Например, хлороглея сарциноидная (Chlorogloea sarcinoides) из синезеленых, в огромных количествах развивающаяся в некоторых соленых озерах, а также ряд других массово растущих водорослей, участвуют в процессе образования лечебных грязей.
Водоросли вневодных местообитаний.
Аэрофильные водоросли.
Аэрофильные водоросли непосредственно контактируют с окружающим их воздухом. Типичное местообитание таких водорослей – поверхность различных внепочвенных твердых субстратов, не оказывающих на поселенцев ясно выраженного физико-химического воздействия (скалы, камни, кора деревьев и т.д.). В зависимости от степени увлажнения, их подразделяют на две группы: воздушные водоросли , обитающие в условиях только атмосферного увлажнения и, следовательно, испытывающие постоянную смену увлажнения и высыхания; и водно-воздушные водоросли , подвергающиеся постоянному орошению водой (брызгами водопада, прибоя и т.д.).
Условия существования водорослей этих сообществ очень своеобразны и характеризуются, прежде всего, частой и резкой сменой температуры и влажности. Днем аэрофильные водоросли сильно прогреваются, ночью охлаждаются, зимой промерзают. Смене условий увлажнения особенно подвержены воздушные водоросли, поскольку они часто вынуждены переходить из состояния избыточного увлажнения (например, после ливня) в состояние минимальной влажности (в засушливые периоды), когда они высыхают настолько, что могут быть растерты в порошок. Водно-воздушные водоросли живут в условиях относительно постоянного увлажнения, однако и они испытывают значительное колебание этого фактора. Например, водоросли, живущие на скалах, орошаемых брызгами водопадов, в летнее время, когда сток существенно уменьшается, испытывают дефицит влаги.
К таким неблагоприятным условиям существования приспособилось сравнительно немного видов (ок. 300). Аэрофильные водоросли представлены микроскопическими водорослями из отделов синезеленых, зеленых и, в значительно меньшей степени, диатомовых и красных водорослей.
При развитии аэрофильных водорослей в массовом количестве они обычно имеют вид порошкообразных или слизистых налетов, войлокообразных масс, мягких или твердых пленок или корочек. Особенно обильны разрастания водорослей на поверхности влажных скал. Они образуют пленки и наросты различного цвета. Как правило, здесь обитают виды, снабженные толстыми слизистыми обвертками. В зависимости от интенсивности освещения, слизь бывает окрашена более или менее интенсивно, что определяет цвет разрастаний. Они могут быть ярко-зеленые, золотистые, бурые, охристые, лиловые, коричневые или почти черные в зависимости от образующих их видов.
Таким образом, аэрофильные сообщества водорослей очень разнообразны и возникают как при вполне благоприятных, так и в экстремальных условиях. Их внешние и внутренние приспособления к такому образу жизни разнообразны и сходны с обнаруживаемыми у почвенных водорослей, особенно развивающихся на поверхности почвы.
Эдафофильные водоросли.
Основной жизненной средой эдафофильных водорослей является почва. Типичные их местообитания – поверхность и толща почвенного слоя, оказывающая на водоросли определенное физико-химическое воздействие. В зависимости от местонахождения водорослей и их образа жизни в пределах этого типа различают три группы сообществ. Это наземные водоросли , массово развивающиеся на поверхности почвы в условиях атмосферного увлажнения; водно-наземные водоросли , массово разрастающиеся на поверхности почвы, постоянно пропитанной водой (в эту группу включаются и водоросли пещер) и почвенные водоросли , населяющие почвенную толщу. Типичные условия – жизнь среди почвенных частиц под влиянием среды, очень сложной по комплексу факторов.
Почва как биотоп имеет сходство с водными и воздушными местообитаниями: в ней есть воздух, причем он насыщен водяными парами, что обеспечивает дыхание атмосферным воздухом без угрозы высыхания. Однако почва кардинально отличается от вышеназванных биотопов своей непрозрачностью. Этот фактор оказывает решающее воздействие на развитие водорослей. Интенсивное развитие водорослей как фототрофных организмов возможно только там, куда проникает свет. В целинных почвах это поверхностный слой почвы толщиной до 1 см, однако в таких почвах водоросли встречаются и на гораздо большей глубине (до 2 м). Это объясняется способностью некоторых водорослей в темноте переходить к гетеротрофному питанию. Многие водоросли сохраняются в почве в состоянии покоя.
Для выживания почвенные водоросли должны иметь способность переносить неустойчивую влажность, резкие колебания температуры и сильную инсоляцию. Эти свойства обеспечиваются у них рядом морфологических и физиологических особенностей (более мелкие размеры по сравнению с водными формами этих же видов, обильное образование слизи). О поразительной жизнеспособности этих водорослей говорит следующее наблюдение: когда почвенные водоросли, хранящиеся десятки лет в воздушно-сухом состоянии в почвенных образцах поместили в питательную среду, они начали развиваться. Почвенные водоросли (преимущественно синезеленые) обладают устойчивостью против ультрафиолетового и радиоактивного излучения.
Характерной чертой почвенных водорослей является способность быстро переходить из состояния покоя к активной жизнедеятельности и наоборот. Они также способны переносить разные колебания температуры почвы. Диапазон выживаемости ряда видов лежит в пределах от –20° до +84° С. Известно, что наземные водоросли составляют значительную часть растительности Антарктиды. Они окрашены почти в черный цвет, поэтому температура их тела оказывается выше температуры окружающей среды. Почвенные водоросли являются также важными компонентами биоценозов аридной зоны, где почва в летнее время нагревается до 60–80° С.
Перечисленные свойства почвенных водорослей позволяют им обитать в самых неблагоприятных местообитаниях. Этим объясняется их широкое распространение и быстрота разрастаний даже при кратковременном появлении необходимых условий.
Подавляющее большинство почвенных водорослей – микроскопические формы, однако нередко их можно увидеть на поверхности почвы невооруженным глазом. Массовое развитие микроскопических форм вызывает позеленение склонов оврагов и обочин лесных дорог, «цветение» пахотных почв.
Число всех видов почвенных водорослей приближается к 2000. Они представлены синезелеными, зелеными, диатомовыми и желтозелеными водорослями.
Литофильные водоросли.
Основной жизненной средой литофильных водорослей служит окружающих их непрозрачный плотный известковый субстрат. Как правило, они обитают в глубине твердых пород определенного химического состава, окруженных воздухом (т.е. вне воды) или погруженных в воду. Различают две группы литофильных сообществ: сверлящие водоросли и туфообразующие водоросли.
Сверлящие водоросли – организмы, внедряющиеся внутрь известкового субстрата. Эти водоросли по количеству видов немногочисленны, однако распространены чрезвычайно широко: от холодных вод севера до постоянно теплых вод тропиков. Обитают они как в континентальных, так и в морских водоемах, у поверхности воды и на глубине более 20 м. Поселяются сверлящие водоросли на известковых скалах, камнях, известковых раковинах животных, кораллах, пропитанных известью крупных водорослях и т.д. Все сверлящие водоросли – микроскопические организмы. Поселившись на поверхности известкового субстрата, они постепенно в него внедряются за счет выделения органических кислот, растворяющих находящуюся под ними известь. Внутри субстрата водоросли разрастаются, образуя при этом многочисленные каналы, при помощи которых они сохраняют связь с наружной средой.
Туфообразующие водоросли – организмы, отлагающие вокруг своего тела известь и обитающие в периферических слоях отлагаемой ими среды, в пределах, доступных для диффузии света и воды. Количество выделяемой водорослями извести различно. Некоторые виды выделяют ее в очень небольших количествах, в виде мелких кристаллов она располагается между особями или образует футляры вокруг клеток и нитей. Другие виды выделяют известь настолько обильно, что постепенно оказываются совершенно погруженными в отложения, что, в конце концов, приводит к их гибели.
Туфообразующие водоросли встречаются в воде и в наземных местообитаниях, в морях и пресных водоемах, в холодных и горячих водах.
Сожительство водорослей с другими организмами
Особый интерес представляют случаи сожительства водорослей с другими организмами. Чаще всего водоросли используют живые организмы как субстрат, наравне с камнями, бетонными и деревянными сооружениями и т.п. По характеру субстрата, на котором поселяются водоросли обрастаний, среди них выделяют эпифиты , поселяющиеся на растениях, и эпизоиты , живущие на животных.
Водоросли могут жить также в тканях других организмов: как внеклеточно (в слизи, межклеточниках водорослей, в оболочках мертвых клеток), так и внутриклеточно. Такие водоросли называются эндофитами . Для них характерно наличие более или менее постоянных и прочных связей между партнерами. Эндофитами могут быть самые разнообразные водоросли, но наиболее многочисленны эндосимбиозы одноклеточных зеленых и желтозеленых водорослей с одноклеточными животными.
Среди симбиозов, образуемых водорослями, наибольший интерес представляет их симбиоз с грибами, известный под названием лишайникового симбиоза , в результате которого возникла своеобразная группа растительных организмов, получившая название «лишайники». Этот симбиоз показывает уникальное биологическое единство, которое привело к появлению принципиально нового организма. Вместе с тем каждый партнер лишайникового симбиоза сохраняет черты той группы организмов, к которой он относится. Лишайники представляют единственный доказанный случай возникновения нового организма в результате симбиоза двух.
Водоросли играют огромную роль в природе. Они являются основными производителями органической пищи и кислорода в водных экосистемах Земли, и, кроме того, играют большую роль в общем балансе кислорода на планете. В наземных местообитаниях почвенным водорослям наряду с другими микроорганизмами принадлежит роль пионеров растительности. Водоросли учувствуют в процессах формирования примитивных почв на субстратах, лишенных почвенного покрова, а также в процессах восстановления почв, нарушенных сильными загрязнениями. Водоросли принимают участие в строительстве коралловых рифов – наиболее грандиозных геологических образований, созданных живыми организмами. Геохимическая роль водорослей, прежде всего, связана с круговоротом в природе кальция и кремния.
Велика историческая роль водорослей. Возникновение кислородосодержащей атмосферы, выход живых существ на сушу и развитие аэробных форм жизни, доминирующих ныне на нашей планете – все это результаты деятельности древнейших фотосинтезирующих организмов – синезеленых водорослей. Массовое развитие водорослей в прошлые геологические эпохи привело к образованию мощных толщ горных пород. От водорослей произошли растения, заселившие сушу.
Трудно переоценить значение водорослей и для жизни человека. Водорослям отводят важную роль в решении ряда глобальных проблем, волнующих все человечество, в том числе продовольственной, энергетической, охраны окружающей среды, освоения недр Земли и богатств Мирового океана, изыскания новых источников промышленного сырья, строительных материалов, фармацевтических препаратов, биологически активных веществ и новых объектов биотехнологии.
Наталья Новоселова



Примерно до середины XX века все растения делили на низшие и высшие. К низшим растениям относили бактерии, грибы, слизевики, водоросли, лишайники . Представители этих групп чрезвычайно разнородны, однако им характерны некоторые общие признаки: отсутствие тканей и дифференциации тела на корень, стебель, лист (т.е. отсутствие органов).
В настоящее время бактерии и грибы выделены в самостоятельные царства живых организмов, а слизевики и лишайники рассматриваются как отдельные группы (отделы) в царстве грибов .
Растениям характерны следующие признаки:
– аэробный фотосинтез;
– наличие в клетках хлоропластов;
– запасное вещество – крахмал;
– плотная целлюлозная клеточная оболочка;
– относительная неподвижность.
По одной из классификаций царство Растения делят на три подцарства: Багрянки (красные водоросли), Настоящие водоросли и Высшие растения.
Вегетативное тело багрянок и настоящих водорослей не расчленено на органы и ткани. Их нередко, по-старому, называют низшими растениями. Однако сейчас багрянки и настоящие водоросли относят к царству Протисты .
Высшие растения , в отличие от низших, – сложные, дифференцированные на органы и ткани многоклеточные организмы, приспособленные к обитанию в наземной среде.
Водоросли – сборная группа преимущественно водных фотосинтезирующих протистов. Возникли в протерозое ~800–900 млн. лет назад.
Понятие «водоросли» в научном отношении страдает большой неопределенностью. Само слово «водоросли» означает лишь то, что это – простейшие организмы, живущие в воде. Однако в воде могут встречаться и семенные (кувшинка, ряска ), высшие споровые (мох фонтиналис , хвощ речной , полушник озерный, папоротник сальвиния ) вторичноводные растения. К тому же в водоемах обитают и фотосинтезирующие бактерии (цианобактерии ), которые не являются растениями или протистами, однако они способны к аэробному фотосинтезу (часто их называют сине-зелеными водорослями). С другой стороны, значительное количество микроскопических водорослей произрастает и на суше (в отличие от «водных» водорослей эти «сухопутные» водоросли легко переносят высыхание и очень быстро оживают при малейшем увлажнении).
Известно более 40 000 видов водорослей, которые ранее объединялись в 9 отделов: красные, диатомовые, зеленые, бурые, пирофитовые, желтозеленые, золотистые, харовые, эвгленовые. В настоящее время вопрос о количестве и составе отделов водорослей, как представителей царства Протисты, окончательно не решен. Общим для водорослей является их способность к автотрофному способу питания благодаря наличию фотосинтезирующего аппарата (вместе с тем у некоторых водорослей наряду с автотрофным существует и гетеротрофное питание). Разные группы водорослей различаются набором пигментов, строением хлоропластов, продуктами фотосинтеза, числом и строением жгутиков. Считается, что отделы водорослей произошли от разных групп одноклеточных организмов, т.е. непосредственно не родственны друг другу. От них, вероятно, происходят наземные хлорофиллоносные растения.
Наука, изучающая водоросли – альгология (от лат. algae – водоросли).
Строение клетки . Организация клетки большинства водорослей мало отличается от организации типичных клеток растений, однако имеет и свои особенности (рис. 4.7.).
Клетка большинства водорослей покрыта плотной оболочкой, состоящей в основном из целлюлозы и пектиновых веществ.
Оболочка водорослей слоистая (неоднородная, 2-х – 3-хслойная). Как правило, внутренние слои оказываются целлюлозными, а наружный – пектиновым, который защищает клетку от губительного действия кислот и других реагентов.
У многих водорослей в оболочке откладываются добавочные компоненты: карбонат кальция (харовые ), альгиновая кислота (бурые ), железо (красные, вольвоксовые ). У диатомовых вместо целлюлозы в клеточной оболочке содержится кремний (он укрепляет матрикс оболочки, создавая структуру, похожую на панцирь).
 Лишь
немногие водоросли являются «голыми»
(т.е. без оболочки, окруженные лишь
плазмолеммой), чаще они покрыты пелликулой
–
плотным
эластичным белковым слоем (эвгленовые
)
и способны
легко изменять форму своего тела.
Лишь
немногие водоросли являются «голыми»
(т.е. без оболочки, окруженные лишь
плазмолеммой), чаще они покрыты пелликулой
–
плотным
эластичным белковым слоем (эвгленовые
)
и способны
легко изменять форму своего тела.
С
Рис. 4.7.
Строение
клетки водорослей.
А – хламидомонада
; Б –спирогира
(часть нити).
Под оболочкой находится протопласт, включающий цитоплазму и ядро (ядра). У большинства водорослей цитоплазма расположена постенным тонким слоем, окружая большую центральную вакуоль с клеточным соком.
У большинства водорослей в клетке имеется всего 1 ядро, но, например, у кладофоры их несколько десятков, а у водяной сеточки (гидродиктион ) – несколько сотен. Клетки с большим количеством ядер называются ценоцитными .
В клетках водорослей присутствуют все органоиды типичные растительным клеткам: эндоплазматическая сеть, рибосомы, комплекс Гольджи, митохондрии, хлоропласты и другие.
Однако, хлоропласты водорослей (хроматофоры ) отличаются от хлоропластов растений огромным разнообразием форм, месторасположением в клетке, а также набором пигментов.
По форме они могут быть чашевидными (хламидомонада ), спиральными (спирогира ), пластинчатыми (мелозира ), цилиндрическими (улотрикс ), звездчатые (зигнема ) и др. (рис. 4.7., 4.8.).
В хлоропластах
имеются
различные пигменты:
хлорофиллы
a,
b,
c,
d;
каротиноиды
(оранжевые), фукоксантин
(бурый), фикоцианин
(синий), фикоэритрин
(красный).
хлоропластах
имеются
различные пигменты:
хлорофиллы
a,
b,
c,
d;
каротиноиды
(оранжевые), фукоксантин
(бурый), фикоцианин
(синий), фикоэритрин
(красный).
В
Рис. 4.8.
Некоторые
формы
хлоропластов
водорослей.
1 – звездчатый у зигнемы
; 2 –
цилиндрический уулотрикса
; 3 –
пластинчатый умелозиры
.
У одноклеточных водорослей присутствует также светочувствительный красный глазок – стигма (улавливает и преобразует свет, необходим для ориентации в пространстве), пульсирующие вакуоли (удаляют из клетки излишки воды, выполняют экскреторную функцию) и жгутики (служат для передвижения). Почти все водоросли, за исключением красных , могут образовывать подвижные клетки.
Строение тела. Водоросли могут быть одноклеточными (хламидомонада, хлорелла), колониальными (водяная сеточка, вольвокс, пандорина) и многоклеточными (нитчатые – спирогира, кладофора ; пластинчатые – фукус, ламинария ; харофитные – хара, нителла). Их размеры колеблются от микроскопических (1 мкм – хлорелла ) до гигантских (до 60 м в длину – макроцистис гигантский ).
Тело многоклеточных водорослей представлено талломом, или слоевищем, не имеет настоящих тканей и органов (листьев, стеблей и корней), хотя у некоторых и есть внешне похожие части (рис. 4.9.).
Слоевище водорослей отличается большим разнообразием морфологических структур, отражающие основные этапы их эволюции. Выделяют следующие основные морфологические структуры водорослей:
– амебоидная – это первичная и наиболее примитивная морфологическая структура свойственна одноклеточным организмам и характеризуется отсутствием постоянной формы клетки, оболочки и жгутиков. Передвигаются эти водоросли, как и амебы, с помощью ложноножек (золотистые и желто-зеленые );
– монадная структура – свойственна одноклеточным организмам и характеризуется наличием у таких клеток одного, двух или нескольких жгутиков, благодаря которым они передвигаются (пирофитовые, золотистые, эвгленовые ); у более высоко организованных монадное строение имеют клетки, служащие для бесполого и полового размножения;
– коккоидная – характеризуется неподвижными клетками с плотной клеточной оболочкой, одиночными или соединенными в колонии (зеленые, золотистые, у диатомовых – это единственная структура);
– палъмеллоидная – представляет собой объединение множества коккоидных клеток, погруженных в общую слизь, но не имеющих плазматических связей (зеленые );
– нитчатая – соединение при помощи цитоплазматических тяжей неподвижных клеток в нити, которые могут быть простыми и ветвящимися, свободноживущими и прикрепленными, объединенными часто в слизистые колонии (зеленые, желто-зеленые и др.);
– разнонитчатая
–
это
усложненная нитчатая структура, состоящая
из нитей стелющихся по субстрату и
отходящих от них вертикальных нитей
(зеленые,
золотистые, красные, бурые
);
разнонитчатая
–
это
усложненная нитчатая структура, состоящая
из нитей стелющихся по субстрату и
отходящих от них вертикальных нитей
(зеленые,
золотистые, красные, бурые
);
– пластинчатая – характеризуется многоклеточными слоевищами в форме пластинок, состоящих из одного, двух и более слоев клеток (зеленые, бурые, красные );
– сифональная – слоевище характеризуется отсутствием клеточных перегородок и большим количеством ядер, это в основном морские водоросли (зеленые, жёлто-зеленые )
– харофитная – крупное многоклеточное слоевище линейно-членистого строения, где различают главный «побег», «листья» и ризоиды (харовые ).
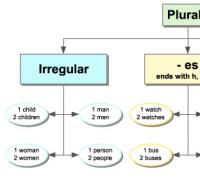
Размножение . Водоросли размножаются бесполым и половым путем.
Бесполое размножение осуществляется вегетативно или спорами.
Вегетативное размножение у одноклеточных осуществляется делением клетки (митоз), у колониальных и нитчатых – частями слоевища, или же специальными органами (например, клубеньками у харовых водорослей).
У некоторых нитчатых водорослей (Spirogira ) нить (тело, таллом) расщепляется строго определенным образом вдоль, и при этом образуются две новые нити – фрагментация .
У ряда нитчатых водорослей (например, улотрикс ) отдельные клетки округляются, накапливают большое количество запасных питательных веществ и пигментов, их оболочки утолщаются. Такие клетки называются акинетами . Они способны переживать неблагоприятные условия, при которых обычные вегетативные клетки водорослей погибают и их нити разрушаются. При наступлении благоприятных условий акинеты прорастают в нити.
Споровое размножение осуществляется при помощи апланоспор (неподвижных спор) или зооспор (подвижных – у большинства водорослей)), образующихся путем деления протопласта обычных клеток или в специальных клетках – спорангиях.
Половое размножение осуществляется с помощью половых клеток – гамет (гаметогамия ), после их слияния образуется зигота, которая дает новую особь или зооспоры. Гаметы образуются в клетках, не отличающихся от вегетативных, или в особых клетках – гаметангиях (женский гаметангий – оогоний, мужской – антеридий). Лишь у харовых водорослей гаметангии многоклеточны. Основные типы полового размножения – изо-, гетеро- и оогамия. Также у водорослей встречаются конъюгация – слияние протопластов неспециализированных неподвижных клеток (спирогира ) и хологамия – слияние двух взрослых одноклеточных подвижных организмов (некоторые вольвоксовые ).
У примитивных водорослей (например, хламидомонада) каждая особь способна формировать и споры, и гаметы в зависимости от времени года и внешних условий (например, температуры). У других функции бесполого и полового размножения выполняют разные особи – спорофиты (образуют споры) и гаметофиты (образуют гаметы). У целого ряда водорослей происходит строгое чередование поколений – гаплоидного гаметофита и диплоидного спорофита.
Экологические группы. Большинство водорослей живет в пресноводных водоемах и морях. Однако существуют и наземные, почвенные и другие экологические группы водорослей.
Выделяют следующие экологические группы водорослей:
– водные : – фитопланктон – совокупность преимущественно мелких, пассивно плавающих в толще воды водорослей (вольвокс, пандорина, фрагилярия, эвглена, хламидомонада ); – фитобентос – совокупность водорослей, живущих на дне водоемов или обрастающие различные водные предметы, а также плавающие на поверхности воды зеленые ватообразные скопления, называемые тиной (хара, нителла, кладофора, улотрикс, спирогира, пиннулярия, новикула, ламинария, фукус );
– наземные (аэрофитные, несколько сотен видов) – водоросли, образующие различно-окрашенные налеты и пленки на деревьях (плеврококк, хлорококк, хлорелла, трентеполия ), скалах, сырой земле, крышах и стенах домов (космарий, пиннулярия), на заборах и т.д.;
– почвенные – водоросли, обитающие на поверхности почвы или в ее самых верхних горизонтах (в РБ ~200 видов: диатомовые, зеленые, желто-зеленые : хламидомонада, хлорелла, хлорококк, навикула, пиннулярия );
– криофитон – водоросли льда и снега (~350 различно-окрашенных видов; явление «красного снега» – скопление одноклеточной водоросли хламидомонады снежной ; зеленая окраска снега – рафидонема снежная ; коричневая окраска снега и льда – анцилонема норденшельда ); и др.
Значение.
1. В водоемах являются основным источником органического вещества – продуценты.
По современным оценкам, на долю океана приходится по меньшей мере половина мировой первичной продукции, выражающейся в количестве фиксированного углерода.
Водоросли (в первую очередь фитопланктон) – очень важное звено в водных экосистемах, с них начинается большинство пищевых цепей.
2. Участвуют в биогеохимических циклах веществ.
Обогащают гидросферу и атмосферу кислородом, причем водорослями выделяется в атмосферу ~50% всего кислорода. Фиксируют углекислый газ. Участвуют в круговороте Ca и Si.
3. Участвуют в почвообразовании.
4. Вступают в симбиоз с другими организмами.
5. Участвуют в процессе естественного самоочищения сточных и загрязненных вод, регенерации воздуха в замкнутых системах (при космических полетах, подводном плавании), могут быть использованы для очистки вод загрязненных радионуклидами (хлорелла способна накапливать радионуклиды).
6. Являются индикаторами загрязнения и засоления среды.
7. Применяются человеком в качестве пищевых продуктов (ламинария, алария, ульва, порфира ), удобрений и кормовых добавок для животных, источник витаминов (А, В 1 , В 2 , В 12 , С и D) и биостимуляторов.
Употреблять в пищу можно, по-видимому, почти все водоросли, так как ядовитых форм среди них нет. Например, на Сандвичевых островах из 115 имеющихся видов водорослей местное население употребляет в пищу около 60.
Особенно богата витаминами красная водоросль порфира . Наибольшей известностью как лечебное и профилактическое средство пользуется морская капуста (ламинария сахарная ), применяемая против некоторых желудочно-кишечных расстройств, склероза, зоба, рахита и др. заболеваний.
8. Являются сырьем для промышленности.
Агар-агар (из красных ), альгин (из бурых ) получаемые из водорослей применяются в пищевой (при производстве мармелада, пастилы, мороженого и т.д.; в качестве добавки к хлебу – не так быстро черствеет), бумажной (придает плотность и глянец), фармацевтической промышленности (для изготовления кремов, мазей), в научных исследованиях (твердая среда для культивирования микроорганизмов).
Диатомин (скопления отмерших диатомовых водорослей) используется почти в 50 отраслях промышленности. Их ценное качество – высокая пористость и низкий удельный вес. Большое количество диатоминов идет на приготовление легких кирпичей; применяется в качестве добавки к цементу. Но наиболее широко применяется в качестве фильтрующего материала при производстве масел, жиров, в сахарной и химической промышленности.
Некоторые вещества используются в парфюмерии, для получения клея (альгиновые кислоты) и т.д. Используются для получения органических кислот, спиртов, лаков и др.
Из бурых водорослей получают йод, бром.
9. Вместе с бактериями вызывают «цветение» воды.
«Цветение» наблюдается при достаточно теплой погоде, когда в воде много питательных веществ (такая ситуация очень часто искусственно создается человеком, когда в воду сбрасывают промышленные стоки или же когда в реки и озера попадают удобрения с полей). В результате начинается взрывоподобное размножение первичных продуцентов, и они в нарушение всех законов природы начинают отмирать раньше, чем их успеют съесть. При последующем разложении остатков происходит столь же интенсивное размножение аэробных бактерий (редуцентов) и вода полностью лишается кислорода. Вследствие чего начинают гибнуть рыбы и другие животные и растения. Токсины, образующие при «цветении» воды, в особенности при размножении цианобактерий (сине-зеленых водорослей), усиливают гибель животных.
Водоросли самые древние растения на Земле, обитающие в воде, на почве, на коре деревьев, а также образующие симбиотический организм - лишайник .
Являются начальным звеном в цепях питания, представляя собой пищу для животных, начиная от простейших и кончая млекопитающими. Кроме того, водоросли в процессе фотосинтеза выделяют в воду кислород, что дает возможность животным дышать в воде как в морях и океанах, так и в маленьких прудах и лужах.
В зарослях водорослей находят приют и среду обитания многие беспозвоночные животные и молодь рыб и земноводных.
Для нормального состояния биоценозов водоемов все должно находиться в равновесии - и ресурсы растений, и количество животных. Для поддержания этого равновесия необходимо, чтобы водоемы были экологически чистыми - в них не сбрасывали бы нечистоты, химические отходы, металлический лом, гниющую древесину и негниющие синтетические материалы, так как это приводит к резкому сокращению количества кислорода, повышению кислотности, увеличению численности гнилостных и болезнетворных бактерий. Это неизбежно ведет к гибели растений и животных, болезням человека и появлению на Земле мертвых и зараженных морей, озер, прудов.
Строение
Водоросли - это низшие споровые растения, содержащие в своих клетках хлорофилл и живущие преимуществепно в воде. В морфологическом отношении для водорослей наиболее существенным признаком является отсутствие тела, расчлененного на стебли, листья и корни. Их тело обозначают как слоевище (или таллом). Размножаются они вегетативно или с помощью спор, т. е. относятся к споровым растениям. В физиологическом отношении водоросли резко отличаются от других групп низших растений наличием хлорофилла,благодаря которому они способны ассимилировать углекислый газ, т. е. питаться фотоавтотрофно. В отличие от водорослей бактерии, имеющие зеленую окраску, содержат пигмент, близкий к хлорофиллу, но не тождественный ему.
Водоросли, даже простейшие из них - сине-зеленые, являются первыми организмами, у которых в процессе эволюции появилась способность осуществлять фотосинтез с использованием воды в качестве источника (донора) водорода и выделением свободного кислорода, т.е. процесс, свойственный высшим растениям. Второй особенностью питания водорослей и других фотосинтезирующих растений является способность усваивать азот, серу, фосфор, калий и другие минеральные элементы в виде ионов минеральных солей и использовать их для синтеза таких важных компонентов живой клетки, как аминокислоты, белки, нуклеиновые кислоты, макроэргические соединения, вещества вторичного обмена. Среди водорослей есть виды, которые являются строгими фотосинтетиками (из синезеленых - анабены, некоторые штаммы ностоков; из зеленых - некоторые виды хлорококкум, хламидомоносов).
Многие водоросли в определенных условиях могут легко переключаться с фотоавтотрофного способа питания на ассимиляцию различных органических соединений, т. е. осуществлять гетеро- или фотогетеротрофный (сочетание гетеротрофного и фотоавтотрофного) типы питания.
Основная структурная единица тела водорослей - клетка. Уникальную группу составляют сифоновые водоросли: у них таллом не разделен на клетки, однако в цикле развития имеются одноклеточные стадии.
Многоклеточные формы возникли после того, как клетка проделала длительный и сложный путь развития в качестве самостоятельного организма. Переход от одноклеточного к многоклеточному состоянию сопровождался потерей индивидуальности и связанными с этим, изменениями в структуре и функциях клетки. С возникновением многоклеточности связаны дифференцировка и специализация клеток в талломе, что следует рассматривать как первый шаг на пути становления тканей и органов.
Краткая характеристика отдельных представителей водорослей
Одноклеточные водоросли Хламидомонада. Отдел: Зеленые водоросли
Подвижные формы (два жгутика на переднем конце). Клетка имеет пектиновую оболочку, которая плотно прилегает к протопласту (у более старых особей слегка отстает от него в задней части клетки). Строение протопласта типичное для вольвоксовых: имеется чашевидный хроматофор с одним пиреноидом (округлое тело, богатое белками, нередко в середине находится белковый кристалл), окруженным крахмалом, ядро, глазок, пульсирующие вакуоли. Способ размножения бесполый - особь теряет жгутики, протопласт внутри материнской оболочки делится последовательно на 2-4 (8) частей. Отрастают два жгутика и особи (зооспоры) выходят наружу. Половое размножение связано с возникновением гамет, слияние которых приводит к образованию зиготы. Гаметы сливаются парами. У большинства видов наблюдается изогамия, но встречаются гетерогамия и настоящая оогамия. Обитатель мелких, хорошо прогреваемых и сильно загрязненных водоемов. Активные санитары загрязненных вод. Хлорелла. Отдел: Зеленые водоросли.
Клетки шарообразные, одноядерные, с чашевидным пристеночным хроматофором с пиреноидом. Клетка покрыта твердой оболочкой целлюлозной природы. Химический состав: белки - 40 % (на сухую массу) и больше, липиды - до 20 % (в среднем), углеводы - до 35 %, зольные вещества - 10 %. Есть витамины С, К и группы В. Обнаружено вещество, имеющее антибиотическую активность, - хлореллин. Размножение бесполое - в материнской клетке образуется около десятка апланоспор (автоспор), которые освобождаются через разрыв ее оболочки. Апланоспоры не имеют жгутиков и еще внутри материнской клетки покрываются целлюлозной оболочкой. Распространена в пресных водоемах, на влажной земле, стволах деревьев, встречается как симбионт с животными (инфузориями, гидрами, червями) - Zoochlorella - и грибами, как гонидий лишайников. Культивируется в искусственных условиях. Нитчатые водоросли Улотрикс. Отдел: Зеленые водоросли.
Тело нитчатое, неразветвленное, прикреплено к субстрату с помощью клетки, вытягивающейся в короткий бесцветный ризоид. Остальные клетки одинаковые, короткие, часто с толстыми набухающими оболочками. Имеется одно ядро, пластинчатый хроматофор с пиреноидами. Размножение бесполое зооспорами с четырьмя жгутиками. (Имеются макро- и микрозооспоры, отличающиеся только размерами.) Половое воспроизведение изогамное. Гаметы образуются на тех же нитях, что и зооспоры, но их больше и они имеют лишь два жгутика. Зигота переходит в состояние покоя и позже прорастает четырьмя зооспорами. При этом происходит редукционное деление ядра и образуются гаплоидные особи. Распространена в реках, где, прикрепляясь к подводным предметам, образует ярко-зеленые обрастания. Спирогира. Отдел: Зеленые водоросли.
Нити состоят из одинаковых цилиндрических клеток с лентообразным, спирально закрученным хроматофором, пристенным слоем протоплазмы, пиреноидами, окруженными мелкими крахмальными зернами, ядром и вакуолей. Клеточная оболочка целлюлозная, извне окружена слизистым чехлом. Клетки способны к делению (вегетативное размножение), которое происходит ночью. Клетки после деления растут только в длину. Нити могут разрываться на отдельные участки. Половое размножение - конъюгация. Нити сближаются, склеиваются своей слизью. Образуются боковые отростки клеток, которые соединяются. На концах отростков образуется канал, соединяющий две клетки. По этому каналу содержимое одной клетки перетекает в другую, сливаясь в зиготу. Ее прорастание происходит после периода покоя. Распространена в стоячих или медленно текущих пресных водоемах. Слизистые на ощупь нити лежат на дне или поднимаются на поверхность в большом количестве. Образуют основную массу тины прудов. |
Существует огромное разнообразие форм (шаровидные, грушевидные, яйцевидные, веретеновидные, спиралевидные, цилиндрические и др.) и размеров (от нескольких микрометров у синезеленых до нескольких сантиметров у харовых) водорослевой клетки.
Размножение
Различают размножение:
- вегетативное
[показать]
.
Вегетативное - деление особей надвое. Иногда делению предшествует отмирание отдельных клеток (у синезеленых), иногда для вегетативного размножения служат специальные образования: почки на слоевищах сфацелярии из бурых водорослей; одно- или многоклеточные клубеньки у харовых водорослей; акинеты (иногда их называют спорами) - клетки, способные переживать неблагоприятные условия у нитчатых синезеленых. Вегетативное размножение - одна из форм бесполого размножения.
- бесполое
[показать]
.
Бесполое размножение сопровождается делением протопласта клетки на части и выходом продуктов деления из оболочки материнской клетки. Бесполое размножение происходит посредством спор или зооспор (спор со жгутиками). Они образуются в клетках, не отличающихся по форме от других клеток, или в особых клетках - спорангиях, которые могут иметь другую форму и размеры, чем вегетативные. Главное отличие спорангиев от других клеток заключается в том, что они возникают как выросты обычных клеток и выполняют только функцию образования спор.
Типы спор:
- апланоспоры - споры, одевающиеся оболочкой внутри материнской клетки;
- автоспоры - апланоспоры, которые в материнской клетке приобретают подобную ей форму.
По количеству их в спорангиях различают тетраспоры (многие красные и диктиотовые из бурых), биоспоры (кораллиновые из красных) и моноспоры (некоторые красные).
Споры и зооспоры обычно выходят в воду через отверстие в стенке спорангия целой группой, окруженные слизистой оболочкой, которая вскоре расплывается.
- половое
[показать]
.
Половое размножение заключается в слиянии двух клеток (гамет), в результате чего образуется зигота, вырастающая в новую особь или дающая зооспоры.
Типы полового размножения:
- соединение содержимого двух вегетативных клеток (гологамия - слияние двух особей у вольвоксовых; конъюгация - слияние содержимого двух безжгутиковых вегетативных клеток у конъюгатов из зеленых водорослей);
- образование внутри клеток специализированных половых клеток-гамет (мужские гаметы имеют жгутики, женские - не всегда). Вместилища гамет называются гаметангиями.
В зависимости от относительных размеров гамет различают:
- изогамию - гаметы одинаковой величины и формы;
- гетерогамию (анизогамию) - женская гамета крупнее мужской, но сходна с нею;
- оогамию - женская гамета (яйцеклетка) лишена жгутиков, неподвижна, значительно крупнее мужской, которую называют сперматозоидом или антерозоидом; гаметангии, содержащие яйцеклетку, называются оогониями, мужскую гамету - сперматангиями или антеридиями;
- автогамию - особый тип полового процесса (у части диатомовых), заключающийся в том, что ядро клетки предварительно делится с мейозом на 4 ядра, два из них разрушаются, а оставшиеся два сливаются, вновь образуя диплоидное ядро. Автогамия не сопровождается увеличением числа особей, а лишь их омоложением.
В результате слияния гамет образуется зигота, жгутики отпадают, появляется оболочка (если жгутики некоторое время сохраняются, зигота называется планозиготой). В зиготе происходит слияние двух ядер - она диплоидна. В дальнейшем зиготы различных водорослей ведут себя по-разному: одни покрываются толстой оболочкой и впадают в период покоя, длящийся до несколььих месяцев; другие прорастают без периода покоя. В одних случаях из зиготы вырастают новые слоевища, в других - из зиготы образуются зооспоры.
Есть водоросли, у которых органы бесполого и полового размножения развиваются на разных особях; тогда растения, образующие споры, называют спорофитами, а растения, производящие гаметы, - гаметофитами. У других водорослей споры и гаметы образуются на одних и тех же растениях.
Распространение в природе
По условиям существования водоросли можно разделить на две группы: живущие в воде и живущие вне воды.
Водные организмы делятся на планктонные (находятся во взвешенном состоянии в воде и имеют ряд приспособлений к этим условиям жизни), бентические (находятся на дне водоемов), перифитонные (обрастают подводные скалы, высшие водные растения, подводные предметы), нейстонные (плавают в полупогруженном состоянии на поверхности воды на границе раздела вода - воздух). Водоросли, живущие вне воды, разделяются на аэрофитон (наземные обрастания) и почвенные.
Кроме названных групп, различают водоросли горячих источников (типичными обитателями их являются синезеленые, специфически термофильных форм мало - мастигокладус, формидиум); водоросли снега и льда (явление "красного снега" вызывает хламидомонада снежная; 80 видов "ледовых" диатомей); водоросли соленых водоемов (дуналиелла солевая из вольвоксовых, хлороглея сарциноидная из синезеленых); водоросли в известняковом субстрате (сверлящие и туфообразующие водоросли - гиелла, ривулярия).
Отделы водорослей
Разделение водорослей на систематические группы - отделы - в основном совпадает с характером их окраски, связанной, конечно, с особенностями строения. Наиболее широко распространено деление водорослей на 10 отделов:
- синезеленые
[показать]
.
Синезеленые водоросли окрашены в синевато-зеленый, иногда почти черно-зеленый или оливково-зеленый цвет. Пигменты: хлорофилл а, каротиноиды, синий фикоцианин, в небольшом количестве красный фикоэритрин. Формы преимущественно многоклеточные, колониальные или нитчатые, есть одноклеточные. Оболочки состоят из муреина, пектиновых веществ, иногда целлюлозы, ослизняются. Характерное строение клеток: нет дифференцированного ядра, хлоропластов, вакуолей, в цитоплазме расположены фотосинтетические мембраны, пигменты и нуклеопротеиды - главная составная часть ядер других растений. У многих синезеленых водорослей в цитоплазме сеть газовые вакуоли, у значительного количества нитчатых образуются так называемые гетероцисты - клетки со специфической структурой.
Размножение у одноклеточных происходит делением клеток, у колониальных и нитчатых - распадом колоний и нитей, есть половой процесс. У многих синезеленых образуются споры для перенесения неблагоприятных условий и для размножения.
- пирофитовые
[показать]
.
Пирофитовые водоросли - одноклеточные, существенным признаком является спинно-брюшное (дорсовентральное) строение их клеток (отчетливо выражены спинная, брюшная и боковые стороны, передний и задний концы). Характерно наличие бороздок, их может быть две (продольная и поперечная) или одна (продольная). Имеются два разных по длине жгутика, глотка (имеющая вид мешка, трубы, внутреннего кармана или треугольного резервуара), сильно преломляющие свет тельца - трихоцисты (располагаются в периферическом слое цитоплазмы, на внутренней поверхности глотки или внутри протопласта). Окрашены обычно в оливковый, бурый или коричневый цвет, нередко желтый, золотистый, красный, реже голубой, синий. Пигменты: хлорофиллы а и с, ксантофиллы, перидинины. Бывают бесцветные формы. Питание автотрофное, реже гетеротрофное. Размножение в основном вегетативное, реже бесполое (зоо- и автоспорами). Половой процесс неизвестен.
Широко распространены в водоемах нашей планеты (пресных, солоноватых водах, в морях).
- золотистые
[показать]
.
Золотистые водоросли - преимущественно микроскопические, одноклеточные, колониальные и многоклеточные формы. Окрашены в золотисто-желтый цвет. Пигменты: хлорофиллы а и с, каротиноиды, особенно много фукоксантина. Живут преимущественно в чистых пресных водах, характерны для кислых вод сфагновых болот. Некоторые виды обитают в морях. Развиваются обычно ранней весной, поздней осенью и зимой.
Строение клетки однотипно: в протопласте находится один или два постенных корытовидных хлоропласта с пиреноидом, ядро небольшое, у некоторых видов на передней части клетки имеется одна или две пульсирующие вакуоли. Оболочка: у простейших представителей - нежный перипласт; у большинства золотистых - перипласт плотный, клетка имеет постоянную форму; у высокоорганизованных представителей - настоящая целлюлозная, обычно двухконтурная оболочка. У многих видов клетки покрыты панцирем с шипами и иглами.
Размножаются простым делением клетки или распадом таллома на части. Наблюдается бесполое размножение с помощью зооспор, реже автоспор. Известен половой процесс в виде типичной изогамии, автогамии.
- диатомовые
[показать]
.
Диатомовые водоросли - микроскопические одноклеточные, колониальные или нитчатые водоросли буровато-желтого цвета. Пигменты: хлорофиллы а и с, каротин, фукоксантин и другие ксантофиллы. Форма клеток разнообразна. Оболочка представлена панцирем из кремнезема, выстланным изнутри пектиновой прослойкой. В кремневом панцире находятся поры - ареолы. В зависимости от конфигурации клеток и структуры створок различают диатомовые с радиальной и двусторонней симметрией.
У многих диатомовых в створках есть продольная щель (так называемый шов). На его концах и посредине находятся утолщения оболочки, которые называются узелками. Благодаря шву и узелкам клетка движется. Водоросли, которые не имеют шва, не двигаются.
Протоплазма расположена в клетках тонким слоем. В клетке находятся ядро, вакуоль с клеточным соком. Хлоропласты имеют различную форму, их бывает один или несколько.
Размножаются диатомовые делением. Есть у них и половой процесс, обычно связанный с образованием ауксоспор, т. е. "растущих спор", которые сильно разрастаются и затем прорастают в клетки, значительно отличающиеся по размерам от исходных. Ауксоспоры свойственны только диатомовым. Диатомовые могут образовывать покоящиеся споры.
Живут повсюду: в водоемах (пресных и соленых), в болотах, на камнях и скалах, в почвах и на их поверхности, на снегу и льду.
- желтозеленые
[показать]
.
Желтозеленые водоросли - одноклеточные, колониальные, многоклеточные и неклеточные формы. Большинство неподвижных, но есть подвижные формы.
Этот отдел недавно выделен из зеленых водорослей. Желтозеленые водоросли отличаются тем, что два жгутика их зооспор неодинаковы по расположению и строению: один - более длинный, направленный вперед, на оси имеет отростки, второй - гладкий, короткий, направленный назад. Оболочка клеток содержит много пектиновых веществ, не дает реакции на целлюлозу.
Строение клеток желтозеленых водорослей однотипно. В протопласте несколько хлоропластов, имеющих дисковидную, корытовидную, пластинчатую, реже лентовидную, звездчатую форму. Основные пигменты: хлорофиллы а, е, каротины и ксантофиллы. У подвижных форм на переднем конце хлоропласта расположен красный глазок. Ядро одно. У немногих видов имеется пиреноид и в передней части клетки одна или две пульсирующие вакуоли.
Вегетативное размножение происходит делением клеток, распадом колоний или нитей, бесполое размножение - зоо- и автоспорами. Половой процесс известен для небольшого количества родов: изо-, оогамия.
Распространены в планктоне, бентосе пресных водоемов, в морях, почве, в местах с повышенной влажностью.
- бурые
[показать]
.
Бурые водоросли - в большинстве случаев это морские формы. Они многоклеточные, прикрепляются к субстрату. Размеры их варьируют от нескольких миллиметров до нескольких метров, есть виды, достигающие 60 м.
По внешнему виду это разветвленные кустики, пластинки, шнуры, ленточки, некоторые как будто имеют стебель и листья. В клетке одно ядро, хлоропласты бурого цвета, зернистые, чаще их много. Пигменты: хлорофиллы а и с, каротины, много фукоксантинов.
В многорядных слоевищах наблюдается специализация клеток с образованием тканей. В простейшем случае различают кору (интенсивно окрашенные клетки, содержащие хлоропласты) и сердцевину (бесцветные крупные клетки одинаковой формы). У более сложно организованных (ламинариевых и фукусовых) различают еще поверхностный слой делящихся клеток, способных производить органы размножения и называемых меристодермой, и промежуточный слой между сердцевиной и корой. Сердцевина служит для транспорта продуктов фотосинтеза и выполняет механическую функцию.
- красные
[показать]
.
Красные водоросли (багрянки) - обширная группа среди донных морских водорослей. Очень редко встречаются в пресных водоемах (виды батрахоспермума), в наземных обрастаниях (порфиридиум). Окрашены в различные оттенки красного цвета, некоторые имеют желтоватую, оливковую или синевато-зеленую окраску.
Пигменты: хлорофиллы a и d (последний встречается только у красных водорослей), каротины, ксантофиллы, R-фикоэритрин, R-фикоцианин. Почти все красные водоросли многоклеточные, имеют форму нитей, разветвленных нитей, кустиков, у некоторых есть стеблеподобные и листообразные органы. Не бывают таких больших размеров, как бурые. Все прикреплены к субстрату. Клетки покрыты оболочкой, состоящей из двух слоев: внутреннего - целлюлозного и наружного - пектинового. У наиболее простоорганизованных клетки одноядерные, у высокоорганизованных - многоядерные. Имеется один или несколько хлоропластов. Особенностью красных водорослей является наличие у некоторых представителей особых железистых клеток. Клетки, слагающие слоевище, соединяются порами.
Вегетативное размножение наблюдается редко. При бесполом процессе совершенно отсутствуют зооспоры. Половой процесс - оогамия.
Багрянки имеют своеобразное строение оогония (карпогона) и сложные процессы развития зиготы. Подвижные стадии в цикле развития отсутствуют. Зигота, прежде чем дать начало спорофиту, претерпевает сложное развитие, в результате которого образуются споры (карпоспоры), дающие начало спорофиту.
- эвгленовые
[показать]
.
Эвгленовые водоросли - микроскопические организмы. Форма клеток в основном эллипсообразная, веретеновидная. Хлоропласты звездчатой, лентовидной, крупнопластинчатой формы.
Пигменты: хлорофиллы а, b, каротин, ксантофиллы. У некоторых эвгленовых имеется красный пигмент - астаксантин, регулирующий количество света, попадающего на хлоропласты. В условиях интенсивного освещения пигмент скапливается в периферической части клетки и затеняет собой хлоропласты. Клетка при этом окрашивается в красный цвет. Целлюлозной оболочки нет, ее роль играет уплотненный слой цитоплазмы; у некоторых имеется панцирь, не связанный плотно с протопластом. На переднем конце тела - углубление (глотка), от дна которого отходит один или два жгутика. Двигаются эвгленовые за счет изменения формы тела и с помощью жгутика, поворачиваясь одновременно вокруг продольной оси. У живых эвгленовых в передней части клетки находится красное пятно - стигма, выполняющее роль светочувствительного органа. Размножаются продольным делением. Наличие полового процесса не установлено. Обитают главным образом в небольших пресных водоемах, некоторые - в солоноватых.
- зеленые
[показать]
.
Зеленые водоросли - это самый многочисленный отдел (до 20 000 видов). Отличаются чисто зеленым цветом своих слоевищ.
Пигменты: хлорофиллы а и b, каротины и много ксантофиллов. У некоторых видов и на некоторых стадиях развития зеленая окраска может маскироваться красным пигментом гематохромом. Одноклеточные, колониальные и многоклеточные формы. Размеры: от мельчайших одиночных клеток диаметром 1-2 мкм до макроскопических растений, измеряемых в длину десятками сантиметров. У зеленых водорослей встречаются все основные типы бесполого и полового размножения и все основные типы смены форм развития.
- харовые
[показать]
.
Харовые водоросли - своеобразная группа водорослей, по внешнему виду напоминающая высшие растения. Широко распространены в пресноводных прудах и озерах, особенно в жесткой известковой воде; встречаются в морских заливах и в солоноватых континентальных водах. Образуют заросли. Высота их слоевищ обычно составляет 20-30 см, но может достигать 1 и даже 2 м. Имеют вид кустистых нитевидных или стеблевидных зеленых побегов членисто-мутовчатого строения: на основных побегах, условно называемых стеблями, на некотором расстоянии друг от друга располагаются мутовки боковых побегов - условно листья - тоже членистого строения. Места расположения мутовок называются узлами, а участки стебля между ними - междоузлиями. Клетки узлов и междоузлий отличаются: междоузлие - это гигантская вытянутая клетка, не способная к делению; узел состоит из собранных в диск нескольких мелких одноядерных клеток, дифференцирующихся в процессе деления и образующих боковые ответвления и мутовку.
Хлоропласты многочисленны, имеют форму мелких дисковидных телец (напоминают хлорофилловые зерна).
Пигменты: хлорофиллы а и b, каротины, ксантофиллы (подобны зеленым водорослям). Своеобразно строение органов полового размножения, образующихся на вершине большинства члеников - листьев. Женские органы - оогонии - и мужские - антеридии - многоклеточны, развиваются обычно на одном растении (редко бывают двудомные).
Роль водорослей в природе, их хозяйственное значение
Водоросли - один из древнейших организмов, населяющих нашу планету. В прошлые геологические эпохи, как и в настоящее время, водоросли населяли океаны, реки, озера и другие водоемы. Обогатив атмосферу кислородом, они вызвали к жизни разнообразный мир животных и способствовали развитию аэробных бактерии; они явились родоначальниками растений, заселивших сушу, и создали мощные толщи горных пород.
Водоросли, как и высшие растения на суше, - источник органических веществ, продуценты кислорода в водоемах. Вследствие деятельности водорослей (в основном диатомовых, синезеленых и зеленых) образуются горные породы (диатомиты, кремнистые залежи, некоторые известняки). Некоторые водоросли (сверлящие синезеленые), разрушая горные породы, участвуют в образовании первичных почв.
В комплексе с другими организмами (бактериями, грибами) водоросли принимают участие в процессе самоочищения воды.
Однако развиваясь в большом количестве, водоросли (синезеленые, некоторые зеленые, диатомовые, пирофитовые) могут приводить к "цветению воды", во время которого значительное количество организмов оседает на дно, усиливаются процессы гниения, резко уменьшается количество кислорода и повышается концентрация углекислого газа. Это приводит к летнему замору рыб. "Цветение" отрицательно отражается на водоснабжении (забиваются фильтры, вода приобретает неприятный вкус и запах).
В сельском хозяйстве водоросли используются как органические удобрения (азотфиксирующие синезеленые водоросли, морские водоросли, а также масса синезеленых водорослей, которую собирают во время "цветения" водоемов). Водоросли обусловливают образование гумуса, улучшают аэрацию почвы, влияют на ее структуру.
Водоросли являются сырьем для получения ценных органических веществ: спиртов, аммиака, лаков, органических кислот и т. п. (сапропели); йода, брома (бурые водоросли); клея (ламинариевые); агар-агара (красные водоросли, филлофора), каротина, биологически активных веществ. Используются в микробиологической промышленности, космических исследованиях. Для производства бумаги и картона используют кладофору и ризоклоний, которые в большом количестве развиваются в водоемах Западной Сибири. Морские водоросли используют в пищевой промышленности, а также непосредственно в пищу (морская капуста, морской салат, ностоки).
В санитарной гидробиологии водоросли используются как индикаторы, показывающие степень загрязненности воды органическими веществами. Водоросли применяют для очистки промышленных вод.
Насколько красив и удивителен подводный мир, настолько же он и загадочен. До сих пор учеными открываются какие-то совершенно новые, необыкновенные виды животных, исследуются невероятные свойства растений, расширяются области их применения.
Растительный мир океанов, морей, рек, озер и болот не столь многообразен, как наземный, однако тоже неповторим и прекрасен. Постараемся разобраться, что же представляют собой эти удивительные каково строение водорослей и значение их в жизни человека и других живых существ.
Систематическое положение в системе органического мира
По общепринятым меркам водоросли считаются группой низших растений. Они входят в состав империи Клеточные и подцарства Низшие растения. На самом деле такое деление основано именно на особенностях строения данных представителей.
Свое название они получили за то, что способны расти и жить под водой. Латинское название - Algae. Отсюда образовано и имя науки, занимающейся подробным изучением этих организмов, их хозяйственным значением и строением, - альгология.
Классификация водорослей
Современные данные позволяют отнести все имеющиеся сведения о разных видах представителей к десяти отделам. В основу разделения положено строение и жизнедеятельность водорослей.
- Сине-зеленые одноклеточные, или цианобактерии. Представители: цианеи, дробянки, микроцистисы и прочие.
- Диатомовые. К ним относятся пиннулярия, навикула, плевросигма, мелозира, гомфонема, синедра и другие.
- Золотистые. Представители: хризодендрон, хромулина, примнезиум и прочие.
- Порфировые. К ним относится порфира.
- Бурые. цистозейра и другие.
- Желто-зеленые. Сюда входят такие классы, как Ксантоподовые, Ксантококковые, Ксантомонадовые.
- Красные. Грациллярия, анфельция, багрянки.
- Зеленые. Хламидомонада, вольвокс, хлорелла и прочие.
- Эвшеновые. К ним относятся самые примитивные представители зеленых.
- как основной представитель.
Данная классификация не отражает строение водорослей, а только показывает их способность фотосинтезировать на разной глубине, проявляя пигментацию того или иного цвета. То есть окраска растения и есть признак, по которому его относят к тому или иному отделу.

Водоросли: особенности строения
Главная их отличительная черта - это то, что тело не дифференцировано на части. То есть у водорослей нет, как у высших растений, четкого разделения на побег, состоящий из стебля, листьев и цветка, и корневую систему. Строение тела водорослей представлено талломом, или слоевищем.
Кроме того, корневая система также отсутствует. Вместо нее в наличии специальные полупрозрачные тонкие нитевидные отростки, называемые ризоидами. Они выполняют функцию прикрепления к субстрату, действуя при этом словно присоски.
Сам таллом может быть очень разнообразной формы и окраски. Иногда у некоторых представителей сильно напоминает побег высших растений. Таким образом, строение водорослей весьма специфично для каждого отдела, поэтому в дальнейшем будет рассмотрено подробнее на примерах соответствующих представителей.

Типы талломов
Слоевище - главная отличительная черта любой многоклеточной водоросли. Особенности строения этого органа заключаются в том, что таллом может быть разных типов.
- Амебоидный.
- Монадный.
- Капсальный.
- Коккоидный.
- Нитчатый, или трихальный.
- Сарциноидный.
- Ложнотканевый.
- Сифоновый.
- Псевдопаренхиматозный.
Первые три наиболее характерны для колониальных и одноклеточных форм, остальные для более совершенных, многоклеточных, сложных по организации.
Данная классификация лишь приблизительна, поскольку у каждого типа имеются переходные варианты, и тогда отличить один от другого практически невозможно. Грань дифференцировки стирается.
Клетка водорослей, ее строение
Особенность этих растений заключается изначально в строении их клеток. Оно несколько отлично от такового у высших представителей. Можно обозначить несколько основных моментов, по которым клетки выделяются.
- У некоторых особей они содержат специализированные структуры животного происхождения - органеллы передвижения (жгутики).
- Иногда встречается стигма.
- Оболочки не совсем такие, как у обычной растительной клетки. Зачастую они снабжены дополнительными углеводными или липидными слоями.
- Пигменты заключены в специализированный орган - хроматофор.
В остальном строение клетки водорослей подчиняется общим правилам такового у высших растений. Они также имеют:
- ядро и хроматин;
- хлоропласты, хромопласты и другие пигментсодержащие структуры;
- вакуоли с клеточным соком;
- клеточную стенку;
- митохондрии, лизосомы, рибосомы;
- аппарат Гольджи, и прочие элементы.
При этом клеточное строение одноклеточных водорослей соответствует таковому у прокариотических существ. То есть также отсутствуют ядро, хлоропласты, митохондрии и некоторые другие структуры.
Клеточное строение многоклеточных водорослей полностью соответствует таковому у высших наземных растений, за исключением некоторых видовых особенностей.

Отдел Зеленые водоросли: строение
В данный отдел входят следующие виды:
- одноклеточные;
- многоклеточные;
- колониальные.
Всего насчитывается более тринадцати тысяч видов. Главные классы:
- Вольвоксовые.
- Коньюгаты.
- Улотриксовые.
- Сифоновые.
- Протококковые.
Особенности строения одноклеточных заключаются в том, что снаружи клетка зачастую покрывается дополнительной оболочкой, выполняющей функцию своеобразного скелета, - пелликулой. Это позволяет ей быть защищенной от внешних воздействий, держать определенную форму, а также со временем формировать на поверхности красивые и удивительные узоры из ионов металлов и солей.
Как правило, в строение зеленых водорослей одноклеточного типа обязательно входит какой-либо органоид передвижения, чаще всего жгутик на заднем конце тела. Запасное питательное вещество - крахмал, масло или мука. Основные представители: хлорелла, хламидомонада, вольвокс, хлорококкум, протококкус.

Очень интересны такие представители сифоновых, как каулерпа, кодиум, ацетобулярия. Их таллом - это не нитчатый и не пластинчатый тип, а одна гигантская клетка, выполняющая все основные функции жизнедеятельности.
Многоклеточные могут иметь пластинчатое строение или нитчатое. Если речь идет о пластинчатых формах, то часто они бывают многослойными, а не только однослойными. Зачастую строение водорослей такого вида очень напоминает побеги высших наземных растений. Чем больше ветвится таллом, тем сильнее сходство.
Основные представители - следующие классы:
- Улотриксовые - улотрикс, ульва, монострома.
- Сцеплянки, или коньюгаты - зигонема, спирогира, мужоция.
Колониальные формы - особые. Строение зеленых водорослей такого типа заключается в тесном взаимодействии между собой большого скопления одноклеточных представителей, объединенных, как правило, слизью во внешней среде. Основными представителями можно считать вольвоксов, протококковых.
Особенности жизнедеятельности
Основные места обитания - это пресные водоемы и моря, океаны. Часто вызывают так называемое цветение воды, заволакивая собой всю ее поверхность. Хлорелла находит широкое применение в скотоводстве, так как очищает и обогащает кислородом воду, а идет на корм скоту.
Одноклеточные зеленые водоросли могут использоваться в космических аппаратах для выработки кислорода в результате фотосинтеза без изменения своей структуры и гибели. По временному отрезку именно данный отдел - самый древний в истории подводных растений.
Отдел Красные водоросли
Другое название отдела - Багрянки. Оно появилось из-за особенного цвета представителей этой группы растений. Все дело в пигментах. Строение красной водоросли в целом удовлетворяет всем основным особенностям строения низших растений. Они также могут быть одноклеточными и многоклеточными, имеют таллом различного типа. Встречаются как крупные, так и крайне малые представители.
Однако их окраска обусловлена некоторыми особенностями - наряду с хлорофиллом эти водоросли имеют ряд других пигментов:
- каротиноиды;
- фикобилины.
Они маскируют собой главный зеленый пигмент, поэтому цвет растений может варьироваться от желтого до ярко-красного и багряного. Происходит это благодаря поглощению практически всех длин волн видимого света. Основные представители: анфельция, филлофора, грацилярия, порфира и другие.

Значение и образ жизни
Способны обитать в пресных водах, однако большинство - все же морские представители. Строение красной водоросли, а конкретно способность вырабатывать особое вещество агар-агар, позволяет широко использовать ее в быту. Особенно это касается пищевой кондитерской промышленности. Также значительная часть особей используется в медицине и напрямую употребляется людьми в пищу.
Отдел Бурые водоросли: строение
Часто в рамках школьной программы изучения низших растений, разных их отделов учитель просит учеников: "Перечислите особенности строения Ответ будет таким: таллом имеет самое сложное строение из всех известных особей низших растений, внутри слоевища, которое имеет зачастую внушительный размер, располагаются проводящие сосуды; сам таллом имеет многослойное строение, из-за чего напоминает тканевый тип устройства высших наземных растений.
Клетки представителей данных водорослей вырабатывают специальную слизь, поэтому снаружи всегда покрыты своеобразным слоем. Запасными питательными веществами являются:
- углевод ламинарит;
- масла (жиры разного типа);
- спирт маннит.
Вот что нужно говорить, если вас просят: "Перечислите особенности строения бурых водорослей". Их на самом деле много, и они уникальны на фоне других представителей подводных растений.

Использование в хозяйстве и распространение
Бурые водоросли - основной источник органических соединений не только для морских травоядных обитателей, но и для людей, живущих в прибрежной зоне. Употребление их в пищу широко распространено у разных народов мира. Из них изготавливают лекарственные средства, получают муку и минеральные вещества, альгиновые кислоты.
Водоросли — наиболее древняя и разнородная группа организмов. Они обитают в водной среде, почве, на поверх-ности растений и в других местах. Большинство водорослей являются автотрофами , так как содержат хлорофилл и мо-гут использовать солнечный свет, но нередко их зеленая окраска маскируется другими пигментами. Некоторые во-доросли утратили способность к фотосинтезу и перешли на гетеротрофный тип питания.
Особенности строения водорослей . По строению тела различают одноклеточные, колониальные и многоклеточ-ные водоросли. Оно может быть представлено талломом , или слоевищем, и не подразделяется на вегетативные орга-ны. По форме многоклеточного таллома выделяют нитча-тые, пластинчатые и сифоновые водоросли.
Клетки многих водорослей похожи на растительные. У них имеются клеточная стенка, одна крупная или не-сколько мелких вакуолей с клеточным соком, а также хлоропласты , которые называются хроматофорами. В хроматофорах находятся пигментные системы, в состав которых входят хлорофиллы (зеленые пигменты), каротиноиды (желто-оран-жевые пигменты) и фикобиллины (сине-фиолетовые пиг-менты). Их соотношение определяет окраску водоросли.
Форма хроматофоров очень разнообразна. Она может быть пластинчатой, цилиндрической, лентовидной, чаше-видной, звездчатой и т. д. В хроматофорах расположены пиреноиды, вокруг которых откладываются запасные веще-ства в виде крахмала или близкого к нему углевода.
Вегетативные клетки таллома снаружи покрыты твер-дой стенкой, состоящей из целлюлозы и пектиновых веществ. Иногда снаружи клеточная стенка покрыта или инкрустирована кремнеземом. Цитоплазма заполняет всю полость клетки или расположена послойно. В клетке нахо-дится одно или несколько ядер. Кроме крахмала, в качестве запасных продуктов могут накапливаться капельки масла.
Размножение водорослей. Для водорослей характерны, как правило, одноклеточные органы размножения, спороношения и полового размножения.
Размножение у водорослей может происходить тремя способами:
Вегетативным (деление клеток пополам, фрагментами колоний и нитей, специализированными структурами — клубеньками — у харовых);
Собственно бесполым (подвижными зооспорами и неподвижными апланоспорами);
Половым (с участием гамет или без образования га-мет, путем слияния ядер вегетативных клеток).
Собственно бесполое размножение осуществляется с по-мощью зооспор или клеточных образований, возникающих внутри вегетативных клеток или в особых органах (зооспо-рангиях или спорангиях) путем деления их содержимого. Вскоре после выхода в воду через отверстия в стенке спо-рангия зооспоры сбрасывают жгутики, покрываются кле-точной оболочкой и прорастают в новую особь.
Половой процесс возможен в формах :
Изогамии, при которой происходит слияние одинако-вых по размеру и форме подвижных гамет;
Гетерогамии, при которой сливаются подвижные га-меты, имеющие одинаковую форму, но отличающиеся по размерам;
Оогамии, когда сливается неподвижная крупная жен-ская гамета — яйцеклетка с мелким подвижным спермато-зоидом.
У некоторых зеленых водорослей половой процесс осуществляется в форме конъюгации.
У одних водорослей одна и та же особь может образовы-вать гаметы или споры — в зависимости от возраста и условий окружающей среды, у других — функции полового и бесполого размножения выполняют разные особи.
Водоросли, на которых развиваются органы бесполого размножения, называются спорофитами, а водоросли, на которых развиваются половые органы, — гаметофитами. Эти два поколения в цикле развития организма могут силь-но отличаться по структуре или, наоборот, быть внешне по-хожими друг на друга. Строго упорядоченные жизненные циклы, сходные с циклами высших растений, существуют лишь у эволюционно продвинутых видов, таких, как пред-ставители бурых и зеленых водорослей.
Классификация водорослей. Многочисленные виды во-дорослей отличаются между собой анатомическим строени-ем как всей особи, так и отдельных клеток, различием в пигментах и других включениях и т. д. На основании этих признаков водоросли подразделяют на 10 отделов. Рассмот-рим те из них, которые нашли применение в биотехнологии.
Бурые водоросли
Свое название бурые водоросли получили из-за высокого содержания в хроматофорах (помимо хлорофилла) буро-го пигмента фукоксантина. Изучено около 1,5 тыс. видов тих водорослей, которые распространены главным образом и морях и океанах, преимущественно в прибрежном мелководье. Иногда их находят и вдали от берегов. Бурые водо-росли считаются важным компонентом бентоса.
Постоянные скопления бурых водорослей известны в юй части Атлантического океана, которое носит название Саргассово море (соответственно сами эти водоросли называют саргассами). Они не являются бентосными, а облада-ют плавучестью благодаря воздушным пузырькам, за счет которых постоянно дрейфуют. В прибрежной же части они ведут обычный бентосный образ жизни.
Таллом бурых водорослей многоклеточный, часто дости-гнет гигантских размеров (до 30-50 м). У самых развитых, крупных видов таллом пластинчатый, т. е. многослойный, и разделен как бы на «ткани», выполняющие разные функ-ции. Клетки бурых водорослей одноядерные. Многочислен-ные хлоропласты чаще дисковидные. Запасные продукты накапливаются в виде ламинарина (полисахарид), маннита (сахароспирт) и масла. Пектиново-целлюлозные клеточ-ные стенки легко ослизняются. Продолжительность жизни бурых водорослей достигает нескольких лет.
Вегетативное размножение бурых водорослей может осуществляться обрывками таллома. У некоторых видов имеются выводковые почки, которые легко отламываются, и вырастают в новую особь.
Бесполое размножение (отсутствует у фукусовых) про-исходит с помощью многочисленных двухжгутиковых зоо-спор, образующихся в одногнездных (изредка — в многогнездных) зооспорангиях, либо с помощью неподвижных тетраспор, образующихся в одногнездных тетраспорангиях.
Половой процесс у бурых водорослей встречается во всех формах. У простейших — в форме изо- или гетерогамии, у наиболее высокоорганизованных (например, у ламинарии) половой процесс оогамный. Размножение ламинарии про-исходит с помощью спор. Зооспоры у нее заключены в мно-гочисленные спорангии, или мешочки. При выходе спор наружу окружающая вода приобретает мутный оттенок. Постепенно споры разносятся течением и оседают на дно, где прикрепляются к шероховатостям и прорастают. На развив-шихся ростках (гаметофитах) образуются разнополые клет-ки. На одних ростках (мужских) появляются мелкие под-вижные сперматозоиды, на других (женских) — яйцеклетки. Осенью происходит оплодотворение будущей ламинарии. Зигота вскоре прорастает в водоросль (спорофит), дости-гающую через год длины 4-5 м. Этой же осенью из развив-шегося растения выходят споры. Старое слоевище разруша-ется, и на его месте развивается новое, которое к весне сле-дующего года достигает нормальной промысловой длины.
Основные представители: ламинария, фукус, падина, макроцистис, алярия и др.
Красные, или багряные, водоросли
Почти все красные водоросли являются морскими оби-тателями, обычными в бентосе, находящемся на значитель-ной глубине. Лишь немногие из них обитают в пресновод-ных бассейнах и в почве.
Разнообразная окраска этих водорослей объясняется на-личием, помимо хлорофилла , еще двух пигментов: красно-го — фикоэритрина и синего — фикоцианина. От соотно-шения этих пигментов окраска таллома может варьировать от малиново-красной до голубовато-стальной. Благодаря та-кому пигментному составу образуется специфический за-писной продукт — багрянковый крахмал, который от йода приобретает буро-красный цвет.
Клеточные стенки вместе с межклеточным веществом у некоторых видов сильно ослизняются, из-за чего весь тал-лом приобретает слизистую консистенцию. В связи с этим многие красные водоросли используют для получения агар-агара.
Большинство красных водорослей — двудомные орга-низмы. Они размножаются бесполым и прогрессивным по-ловым путем. Многие «багрянки» характеризуются правильной сменой гаметофита и спорофита, внешне неотличи-мых друг от друга. У некоторых красных водорослей циклы развития носят сложный характер.
Основные представители: порфира, филлофора, анфелъция и др.
Зеленые водоросли
Зеленые водоросли характеризуются травянисто-зеленой окраской, зависящей от преобладания хлорофиллов над каротиноидами. Клетка большинства таких водорослей покрыта целлюлозной оболочкой. У многих из них наблю-дается свойственное высшим растениям правильное чередо-вание бесполого и полового поколения; некоторые зеленые водоросли перешли к наземному существованию.
Представители этого отдела (около 15 тыс. видов) рас-пространены в пресных водах, некоторые — в морях, и очень немногие обитают в условиях периодического увлаж-нения (на почве, стволах деревьев, заборах, цветочных горшках и т. д.).
Типичным представителем является водоросль рода Хламидомонада. Это одноклеточная водоросль, со жгутика-ми, многочисленные виды которой обитают в лужах, кана-вах и других мелких пресных водоемах. В случае их массо-вого развития вода нередко принимает зеленую окраску. При подсыхании водоема хламидомонады теряют жгутики, ослизняются и в таком неподвижном состоянии пережида-ют неблагоприятные условия, а при попадании в воду клет-ки снова вырабатывают жгутики и возвращаются к подвиж-ному состоянию. В благоприятных условиях эти водоросли интенсивно размножаются бесполым путем, формируя большое количество зооспор. У большинства видов половой процесс изогамный.
Род Хлорелла широко распространен в пресных водах, где эта водоросль придает воде зеленый цвет. Она встреча-ется также на сырой земле, на коре деревьев и т. д. Хло-релла — представитель одноклеточных зеленых водорос-лей, у которых отсутствуют жгутики. При бесполом разм-ножении содержимое клетки распадается, образуя от 4 до 64 дочерних клеток, которые освобождаются после разрыва стенки материнской клетки. Половой процесс отсутствует. В клетках хлореллы накапливается много запасных про-дуктов, витаминов, антибиотиков, поэтому ее культивиру-ют в различных целях.
Ярким представителем зеленых водорослей, имеющих таллом в виде разветвленной нити, сложенной из одноядер-ных клеток, может служить род Улотрикс. Эти водоросли встречаются в прибойной полосе больших озер, образуя ва-тообразные обрастания на камнях.
К классу конъюгатов относится представитель рода Спирогира, имеющий нитчатый таллом без жгутиков. Мно-гочисленные виды этого рода имеют лентовидные, спираль-но закрученные хроматофоры с пиреноидами, окруженные крахмальными зернами. Ядро находится в центре клетки и погружено в цитоплазму.
Половой процесс — конъюгация — заключается в слия-нии протопластов вегетативных клеток. Это так называе-мая лестничная конъюгация, которая происходит между клетками параллельно расположенных нитей. Образовав-шаяся в результате слияния протопластов конъюгирующих клеток зигота вырабатывает толстую стенку и переходит в состояние покоя. Ядра сливаются незадолго до прорастания зиготы, после чего образуются четыре гаплоидных ядра, причем из четырех ядер только одно остается жизнеспособ-ным и поэтому развивается только одна особь. Помимо конъюгации, широко распространено вегетативное размно-жение. Оно осуществляется благодаря разрыву нитей на отдельные участки, клетки которых начинают делиться и образуют новые нити.
Основные представители: хлорелла, улъва, спирогира, уло- щрикс, вольвокс, эвглена и др.
Диатомовые
Клеточные стенки диатомовых водорослей состоят в основном из кремнезема, образующего защитный панцирь, который имеет две отдельные части — теки: верхнюю — эпитеку и нижнюю — гипотеку. Пояс эпитеки плотно надвинут на поясок гипотеки. В створках теки имеются | кнозные отверстия — поры, обеспечивающие обмен ве-ществ, а также пустоты. Внутри клетки находятся протопласт и вакуоли. Ядро одно. Хлоропласты имеют бурую окраску, так как хлорофилл в них замаскирован бурыми пиг-ментами — каротиноидами и диатомином (пигмент из группы ксантофиллов). Запасные продукты откладываются в виде масла, волютина и лейкозина.
Основные представители: пиннулярия, навикула, мело- щра, табеллярия и др.
Значение водорослей. Водоросли, обитающие в воде, подразделяют на две большие группы: планктонные и бентосные.
Планктоном называют совокупность свободно плаваю-щих в толще воды на небольшой глубине мелких — пре-имущественно микроскопических — организмов. Расти-тельная часть планктона, образуемая водорослями, состав-ляет фитопланктон. Значение фитопланктона для обитателей водоема огромно, так как им производится основная масса органических веществ, т. е. водоросли является продуцентами в цепи питания.
К бентосным водорослям относятся особи, прикрепленные ко дну водоемов, находящиеся в воде на глубине 30-50 м.
Однако наиболее теневыносливые бурые и красные водорос-ли достигают глубины 100-200 м, а отдельные виды — 500 м и более.
Водоросли живут на почве и даже в атмосферном возду-хе (некоторые виды хлореллы). Отдельные виды, попадая вместе с бактериями на бесплодные субстраты, становятся пионерами их заселения. Многие водоросли активно участ-вуют в процессе почвообразования. Азотфиксирующие во-доросли (анабена) накапливают в почве азот. Некоторые ви-ды водорослей (носток и др.) входят в состав комплексных организмов — лишайников.
Хозяйственное значение водорослей заключается в непо-средственном использовании их в качестве пищевых про-дуктов или как сырья для получения различных веществ, ценных для человека.
Из многочисленных видов водорослей съедобными в на-стоящее время считаются 80 (в основном это морские виды — ламинария, порфира, ульва, спируллина и т. д.). Съедоб-ные водоросли богаты минеральными веществами, особенно йодом. Среди красных водорослей порфира считается дели-катесом во многих приморских странах. В Японии насчи-тывается более 300 наименований блюд из морской капус-ты. Одно из самых популярных блюд с водорослями — су-ши. Под общим названием mozuku биологи обнаружили ряд из шести видов водорослей — kombu, wakame, nori, hijiki и др., которые употребляют в пищу. По статистиче-ским данным, только сырых водорослей японцы съедают в год лишь в 35 раз меньше по весу, чем риса, который, как известно, в этой стране считается блюдом номер один.
Одноклеточные водоросли выращивают в условиях мяг-кого теплого климата (Средняя Азия, Крым) в открытых бассейнах на специальной среде. К примеру, за теплый пери-од года (шесть — восемь месяцев) можно получить 50-60 т биомассы хлореллы с 1 га, тогда как одна из самых высо-копродуктивных трав — люцерна дает с той же площади только 15-20 т урожая. Хлорелла содержит около 50 %" белка, а люцерна — лишь 18 %. В целом в пересчете на 1 га хлорелла образует 20-30 т чистого белка, а люцерна — 2- 3,5 т. Кроме того, хлорелла содержит: углеводы — 40 %, жиры — 7-10 %, витамины А (в 20 раз больше), В 2 , К, РР и многие микроэлементы. Варьируя состав питательной среды, можно в клетках хлореллы сдвинуть процессы био-синтеза в сторону накопления либо белков, либо углеводов, а также активировать образование тех или иных витами-нов. В клетках хлореллы содержится также антибиотик хлореллин.
Водоросли служат кормом для рыб и водоплавающих птиц. В ряде стран их используют как витаминную добавку к кормам для сельскохозяйственных животных. Так, во Франции, Шотландии, Швеции, Норвегии, Исландии, Япо-нии, Америке, Дании и на Русском Севере водоросли прибавляют к сену или дают как самостоятельный корм коро-нам, лошадям, овцам, козам, домашней птице. Для этой цели строят заводы. Опыты, проведенные в Мурманской об-ласти России, показали, что водорослями можно заменить примерно 50 % сочных и 30 % грубых кормов в суточном рационе животного. При этом удои молока и яйценоскость у птиц повышались на 10 % и выше.
Водоросли могут служить удобрением. В таком качестве их широко применяют в Ирландии, Шотландии, Норвегии, Франции. Запахивание биомассы водорослей обогащает поч-ну фосфором, калием, йодом и значительным количеством микроэлементов, а также пополняет почвенную азотфиксирующую микрофлору. При этом водоросли разлагаются в почве быстрее, чем навозные удобрения, и не засоряют ее семенами сорняков, личинками вредных насекомых, спора-ми фитопатогенных грибов. Применение водорослевого пе-регноя и запахивание штормовых выбросов на 140-300 % повышает урожайность не только злаковых культур (пше-ницы, ячменя), но и овощей.
В Израиле на опытных установках проводят экспери-менты с зеленой одноклеточной водорослью Dunaliella, ко-торая способна синтезировать глицерол. Dunaliella может расти и размножаться в среде с широким диапазоном со-держания соли: в морской воде и в почти насыщенных рас-творах Мертвого моря. Она накапливает свободный глице-рол как осмопротектор, чтобы тем самым противодейство-вать высоким концентрациям солей в среде выращивания.
При таких условиях выращивания дуналиеллы на долю глицерола приходится до 85 % сухой массы клеток. В ней содержится также значительное количество β-каротина. Та-ким образом, культивируя эту водоросль, можно получать глицерол, пигмент и белок, что весьма перспективно с эко-номической точки зрения.
Красные водоросли (роды: анфельция, гелидиум, грацилярия) служат источником получения агар-агара (желирующего вещества, широко применяемого в кондитерской, бумажной, фармацевтической промышленности и в микро-биологии). Получают агар-агар (далее — агар) длительным кипячением водорослей. После остывания образуется плот-ное желеобразное вещество, которое применяют при изготов-лении мармелада, пастилы, стабилизации многих консервов, сиропов, шоколадных напитков, мороженого. Кожа, бумага или ткань, обработанные агаром, становятся более прочны-ми и приобретают приятный блеск.
У других багрянок (роды: литотамнион, литофиллум) клеточные стенки инкрустированы известью, которая при-дает таллому твердость камня. Такие красные водоросли принимают участие в образовании коралловых рифов.
Зола водорослей служит сырьем для получения брома и йода. Со времен открытия йода (середина XIX в.) Норвегия и Шотландия извлекали его почти исключительно из дон-ных водорослей. Во время Первой мировой войны, когда потребность в препаратах йода резко возросла, японские за-воды, переработав миллионы тонн сырых водорослей, полу-чили около 600 т йода.
Некоторые водоросли служат в качестве индикаторных организмов при определении степени загрязнения водое-мов. Например, массовое развитие осциллятории — показа-тель степени загрязнения при биологическом анализе воды. Применяют водоросли и для биологической очистки сточ-ных вод, а также — благодаря высокой скорости размноже-ния — для получения биомассы, используемой в качестве топлива.
Известны горные породы (диатомиты, горючие сланцы, часть известняков), возникшие в результате жизнедеятельности водорослей в прошлые геологические эпохи. Диатомин применяют в производстве материалов для звуковой и тепловой изоляции, при изготовлении фильтров для пище-вой и химической промышленности, шлифовке металлов. Водоросли участвуют в образовании лечебных грязей.
Бурые водоросли образуют подводные луга с огромной фитомассой. Они приобретают все возрастающее значение как кормовые, пищевые, лекарственные и технические рас-тения. В северных и умеренных широтах произрастает ла-минария — морская капуста, таллом которой достигает в длину 20 м и содержит много незаменимой аминокислоты метионина, йода, углеводов, минеральных веществ и вита-минов. Из ламинарии также получают алъгинит — клея-щее вещество, используемое в текстильной (ткани не вы-цветают и не промокают) и пищевой (при изготовлении консервов, соков) промышленности, при производстве ме-лованной бумаги. Альгинит повышает устойчивость лако-красочных покрытий и строительных материалов. Эту во-доросль культивируют в морях России и стран Юго-Восточ-ной Азии.
Благодаря таким свойствам водорослей, как простота строения, быстрый рост и скорость размножения, их широ-ко применяют в научных исследованиях по молекулярной биологии, генетике, генетической инженерии, биохимии и физиологии.
Предпринимаются попытки использовать некоторые вы-сокопродуктивные и неприхотливые водоросли (например, хлореллу, которая синтезирует белки, жиры, углеводы, ви-тамины и способна поглощать вещества, выделяемые чело-веком и животными) для создания замкнутого круговорота веществ в обитаемых отсеках космических кораблей.
Лишайники
Лишайники — это симбиотические ассоциации микро-скопических грибов и зеленых микроводорослей и/или ци-анобактерий, образующие слоевища (талломы) определен-ной структуры. Они выделяют кислоты и тем самым вносят существенный вклад в процессы почвообразования. Лишайники можно отнести к пионерам, т. е. к первым организ-мам, заселяющим субстрат в процессе первичной сукцессии.
Преимуществом лишайников является устойчивость к экстремальным условиям (засухе, морозам, высоким темпе-ратурам, ультрафиолетовому излучению). В то же время они проявляют повышенную чувствительность к загрязне-нию окружающей среды и могут служить индикаторами ее состояния.
Строение лишайников . Лишайники представляют собой симбиотическую ассоциацию фотосинтезирующего организ-ма, или фотобионта (водоросли или цианобактерии), и гриба (микобионта). Водоросли и цианобактерии питаются автотрофно, но воду и ионы они берут от гриба. Как прави-ло, грибной мицелий служит для водоросли защитной обо-лочкой, предохраняющей ее от высыхания. Сам гриб, не-способный синтезировать органические вещества, питается гетеротрофно ассимилятами партнера по симбиозу. Однако оба партнера могут существовать и как самостоятельные организмы.
По внутреннему строению лишайники подразделяют на:
Гомеомерные, когда клетки водоросли (фотобионта) распределены хаотично среди гиф гриба по всей толщине таллома;
Гетеромерные, когда таллом на поперечном срезе можно четко разделить на слои.
Большинство лишайников имеет гетеромерный таллом. В гетеромерном талломе верхний слой — корковый, сложен-ный гифами гриба. Он защищает таллом от высыхания и ме-ханических воздействий. Следующий от поверхности слой — гонидиальный. В нем располагается фотобионт. В центре имеется сердцевина, состоящая из беспорядочно перепле-тенных гиф гриба. В ней в основном запасается влага. Сердцевина выполняет также роль скелета. У нижней по-верхности таллома часто находится нижняя кора, с по-мощью выростов которой (ризин) лишайник прикрепляется к субстрату.
В образовании лишайников участвует около 20 % из из-вестных видов грибов (из них аскомицены — около 98 %, дейтеромицеты — около 1,6 %, базидиомицеты — около 0,4 %). Из водорослей в лишайниках наиболее распростра-нена Trebuxia. Из цианобактерий часто встречаются Nostoc, (".alotrix. Цианобактерии как симбионты лишайников спо-собны осуществлять фотосинтез и фиксацию атмосферного азота.
По строению тела (таллома, или слоевища) лишайники бывают накипными (корковыми), листоватыми и кустис-тыми. Они распространены по всему земному шару — от тропиков до приполярных областей. Хорошо известны та-кие лишайники, как исландский мох (Cetraria islancLica) и ииды Usnea, свешивающиеся с деревьев наподобие бороды и очень похожие внешне на цветковые эпифитные растения рода Tillandsia.
Размножение. Большинство лишайников способно реге-нерировать даже из мелких фрагментов слоевища, содержа-щих и фотобионт, и микобионт. У многих групп лишайников по краям или на верхней поверхности слоевища образуются особые выросты — изидии, которые легко отламываются и дают начало новому слоевищу. В других случаях клетка фотобионта в сердцевине лишайника окружается несколь-кими слоями гиф, превращаясь в крошечную гранулу, на-зываемую соредией. Каждая соредия способна прорасти в новое слоевище. Хотя бесполое размножение лишайников достаточно эффективно, у грибов, образующих лишайники, широко распространен и половой процесс.
Значение лишайников. Лишайники настолько выносли-вы, что растут даже там, где отсутствует другая раститель-ность, например, в Арктике и Антарктике. Они первыми за-селяют безжизненные субстраты, в частности камни, и на-чинают почвообразовательный процесс, необходимый для освоения этой среды растениями.
Ряд лишайников служит важным кормом для живот-ных (например, ягель, или олений мох (Cladonia rangife- rina), — корм северных оленей). При нехватке другой пи-щи его едят иногда и люди. Определенные виды лишайни-ков считаются в Китае и Японии деликатесами.
Из лишайников можно получать красители, в частности лакмус, экстрагируемый из видов рода Roccella. Лакмус до сих пор широко применяют в химических лабораториях для быстрого и простого определения реакции среды: в кислой среде он краснеет, а в щелочной синеет. Другие лишайни-ковые красители в свое время использовали для окраски шерсти.
Лишайники очень чувствительны к загрязнителям воз-духа, особенно к диоксиду серы (сернистому газу). При этом степень чувствительности варьирует у разных видов, поэтому их используют в качестве биоиндикаторов степени загрязнения окружающей среды.
Находят применение лишайники и в народной медицине, а выделяемые из них лишайниковые кислоты (усниновая кислота и др.) используют в качестве компонента лекарст-венных средств от ряда заболеваний, например кожных.
Из некоторых лишайников (дубовый мох Evernia prunastri и др.) получают душистые вещества, применяе-мые в парфюмерии.
Грибы
Грибы представляют собой обширную группу организ-мов, включающую около 100 тыс. видов. Это гетеротроф-ные организмы, лишенные хлорофилла. Минеральные ве-щества гриб способен усваивать из окружающей среды, од-нако органические вещества он должен получать в готовом виде.
Строение грибов. Вегетативное тело большинства гри-бов — мицелий — представляет собой переплетение тон-ких ветвящихся нитей (гиф). Мицелий бывает неклеточ-ный (лишен перегородок), представляющий собой как бы одну гигантскую клетку с множеством ядер, и клеточный, разделенный на клетки, содержащие одно или много ядер.
Клеточная стенка грибов содержит до 80-90 % полиса-харидов, связанных с белками и липидами. Скелетные ее компоненты состоят из хитина или целлюлозы. Запасные продукты клеток грибов — гликоген, волютин, масло.
Размножение грибов. Грибы размножаются нескольки-ми способами. Бесполое размножение может быть вегета-тивным и собственно бесполым. Под вегетативным размно-жением подразумевают почкование гиф или отдельных клеток (например, у дрожжей). Образующиеся почки посте-пенно отделяются, растут и со временем сами начинают почковаться. Собственно бесполое размножение осуществ-ляется посредством спор и конидий, которые обычно обра-зуются на специальных ветвях мицелия.
В зависимости от способа образования, различают эндо-генные и экзогенные споры. Эндогенные споры характерны для бесполого размножения низших грибов. Они образуют-ся внутри особых клеток, называемых спорангиями. Экзо-генные споры обычно называют конидиями. Они имеются у низших и у некоторых низших грибов. Конидии образуют-ся на вершинах или сбоку специальных гиф — конидиеносцев, ориентированных вертикально. Конидии покрыты плотной оболочкой, поэтому устойчивы, но неподвижны.
При половом размножении для низших грибов свойственно слияние гаплоидных клеток путем изогамии, гетеро-гамии и оогамии с образованием зиготы, которая покрыва-ется толстой оболочкой, некоторое время проводит в со-стоянии покоя, после чего прорастает. В случае оогамии развиваются половые органы — оогонии (женские) и анте-ридии (мужские).
Классификация грибов. Классификация основных отде-лов царства грибов основана на способе их размножения.
Зигомицеты (Zygomycota)
Это грибы с неклеточным мицелием или с небольшим количеством перегородок; у наиболее примитивных — в ви-де голого комочка протоплазмы — амебоида или в виде од-ной клетки с ризоидами.
Основные представители: мукор, ризопус.
Аскомицеты, или сумчатые грибы (Ascomycota)
Это грибы с многоклеточным гаплоидным мицелием, на котором развиваются конидии. Характерно образование су-мок с аскоспорами — основными органами размножения. Аскомицеты представляют собой одну из самых многочис-ленных групп грибов, которая насчитывает более 32 тыс. видов (примерно 30 % всех известных науке видов грибов). Их отличает огромное разнообразие — от микроскопиче-ских почкующихся форм до обладающих очень крупными плодовыми телами грибов.
Основные представители: хлебные дрожжи, пеницилл, аспергилл, спорынья, пецица, сморчок.
Базидиомицеты (Basidiomycota)
Основные представители: белый гриб, шампиньон, мухо-мор и т. д.
Аско- и базидиомицеты часто объединяют в группу выс-ших грибов.
Дейтеромицеты, или несовершенные грибы (Deuteromycota)
В эту гетерогенную группу объединены все грибы с чле-нистыми гифами, но с неизвестным до настоящего времени половым процессом. Насчитывается около 30 тыс. видов несовершенных грибов.
Значение грибов. Съедобные грибы (белые, сыроежки, грузди и др.) употребляют в пищу, но только после обра-ботки. Наиболее ценный гриб — французский черный трю-фель, для него характерен привкус прожаренных семечек или грецких орехов. Этот гриб является деликатесом. Он растет в дубовых и буковых рощах, главным образом в Южной Франции и Северной Италии.
Искусственное выращивание съедобных грибов способно ннести существенный вклад в дело обеспечения продоволь-ствием все увеличивающегося населения земного шара. Не-обходимо сделать съедобные грибы такой же управляемой сельскохозяйственной культурой, как зерновые злаки, ово-щи, фрукты. Наиболее легко поддаются искусственному ныращиванию древоразрушающие грибы.
В пищевой промышленности различные дрожжевые культуры применяют в хлебопечении, для приготовления уксуса и спиртных напитков (вина, водки, пива, кумыса, кефира), а плесневые культуры — для изготовления сыров (рокфор, камамбер), соевого соуса (Aspergillus oryzae), а также некоторых вин (херес).
Грибы и препараты из них широко применяют в меди-цине. Некоторые виды грибов продуцируют важные веще-ства, в том числе антибиотики — пенициллы, стрептомицеты. В списке официальных препаратов содержатся много-численные препараты из грибов, например из чаги, спорыньи. В восточной медицине используют цельные гри-бы — рейши (ганодерма), шиитаке и др.
Многие грибы, способны к взаимодействию с другими организмами посредством своих метаболитов или прямо ин-фицируя их. Применение сельскохозяйственных пестицидных препаратов из некоторых грибов рассматривается как возможность управления размерами популяций вредите-лей сельского хозяйства, таких, как насекомые, нематоды.
В качестве биопестицидов (препарат боверин) использу-ют, например, энтомопатогенные грибы. Мухомор издавна применялся как инсектицид.
Разнообразны и биотехнологические функции грибов. Их используют для получения таких продуктов, как:
Лимонная кислота (аспергиллус);
Гиббереллины и цитокинины (физариум и ботритис);
Каротиноиды (астаксантин, придающий мякоти лосо-севых рыб красно-оранжевый оттенок, вырабатывают гри-бы Rhaffia rhodozima);
Белок (Candida, Saccharomyces lipolitica);
Trichosporon cutaneum, окисляющий многочисленные органические соединения, включая некоторые токсичные (например, фенол), играет важную роль в системах аэроб-ной переработки стоков.
Плесени также продуцируют ферменты, используемые в промышленности (амилазы, пектиназы и т. д.).
Грибы принимают участие в образовании симбиотиче-ской микоризы с корнями высших растений. Гриб получает от дерева органические соединения, а сам делает воду и ми-неральные вещества доступными для поглощения и всасы-вания растением. Кроме того, гриб обеспечивает дерево большей поверхностью всасывания.
Однако некоторые грибы оказывают и отрицательное воздействие. Так, отдельные представители плесневых гри-бов существенно снижают урожай сельскохозяйственных культур. Грибы-древоразрушители вызывают быструю дест-рукцию деревьев и древесных материалов, поэтому рас-сматриваются как патогенные. Известно большое количест-во разнообразных патогенных грибов, вызывающих заболе-вания растений, животных и человека.